| └тҐюЁ | ╤ююсїхэшх |
|
|
| |
╧юёҐ N: 164
╟рЁхушёҐЁшЁютрэ: 24.02.13
╟рьхірэш : 
|
|
╬ҐяЁртыхэю: 05.10.13 02:15. ╟руюыютюъ: ├ряыюуЁґяя√ Y-─═╩ ш ¤ҐэюухэхҐшіхёъшх яЁюІхёё√
╚ч ёҐЁґъҐґЁ√ уряыюуЁґяя Y-─═╩ фрцх яЁш яютхЁєэюёҐэюь шчґіхэшш ьюцэю ёфхы𥹠т√тюф, іҐю фхЁхтю уряыюуЁґяя Y-─═╩ ЄюЁьшЁютрыюё№ яюф тючфхщёҐтшхь ьэюуюішёыхээ√є ьшъЁю ърҐрёҐЁюЄ, яЁштюфштЇшє ъ Ёхуґы Ёэ√ь яЁюєюфрь іхЁхч ¤ЄЄхъҐ сґҐ√ыюіэюую уюЁы√Їър. ╤ яюьюї№■ ҐхюЁшш ярёёшюэрЁэюёҐш ьюцэю юс· ёэшҐ№ яЁюҐхърэшх ьэюушє яЁюІхёёют ЄюЁьшЁґ■їшє фхЁхтю уряыюуЁґяя Y-─═╩. ┬юч№ь╕ь эряЁшьхЁ ёрьюх Ёрээхх іхыютхіхёҐтю, яЁюшчюЇхфЇхх юҐ ╙ єЁюьюёюьэюую └фрьр. ╧ЁюёҐющ ыюушъющ ҐЁґфэю юс· ёэшҐ№ яюіхьґ шч тёхщ яюяґы Ішш, ъръ с√ юэр эх с√ыр ьрыр, сюы№Їґ■ ірёҐ№ яюҐюьёҐтр юёҐртшы юфшэ іхыютхъ, яюыґіштЇшщ ъ Ґюьґ цх ьґҐрІш■ B-M60. ╩ръшь юсЁрчюь юэ ёьюу яЁхґёяхҐ№, тхф№ ґ эхую с√ыш фхё Ґъш, ёюҐэш, р ьюцхҐ с√Ґ№ ш Ґ√ё іш ъюэъґЁхэҐют. ┼ёыш с√ ¤Ґю с√ы хфшэшіэ√щ ёыґірщ, Ґръ тхф№ уряыюуЁґяяр ┬ шьххҐ эрсюЁ ьґҐрІшщ (M60, M181, P85, P90) ╥ю хёҐ№ Ґръшє яЁюєюфют іхЁхч сґҐ√ыюіэюх уюЁы√Їъю с√ыю іхҐ√Ёх, яЁхцфх, іхь яЁюшчюЇыю фры№эхщЇхх тхҐтыхэшх тэґҐЁш ¤Ґющ тхҐтш. | ІшҐрҐр: | | ├ряыюуЁґяяр CT. ╤юёҐрты ■їшх ¤Ґґ уЁґяяґ ьґцішэ√ юсырфр■Ґ Y-─═╩ ё ╬═╧ ьґҐрІш M168 тьхёҐх ё P9.1 ш M294. ▌Ґш ьґҐрІшш яЁшёґҐёҐтґ■Ґ ґ тёхє ёютЁхьхээ√є іхыютхіхёъшє ьґцёъшє ышэшщ ъЁюьх A ш B, ъюҐюЁ√х яюіҐш яюыэюёҐ№■ юсх эрєюф Ґё т └ЄЁшъх. |
|
╚ юя Ґ№ ґЇы√щ ьґцшіюъ, юёэютртЇшщ тхҐт№ ╤╥, юя Ґ№ ҐЁш ьґҐрІшш, ҐЁш сґҐ√ыюіэ√є уюЁы√Їър, яЁхцфх, іхь хую яюҐюьёҐтю эрірыю тхҐтшҐ№ё . ┬ё╕ ¤Ґю ёыхф√ фЁхтэшє яЁюІхёёют ¤Ґэюухэхчр. ┬ ¤яшІхэҐЁх ¤Ґэюухэхчр т Ёхчґы№ҐрҐх тэхЇэхую тючфхщёҐтш т яюяґы Ішш тючэшър■Ґ ьґҐрІшш яЁштюф їшх эх Ґюы№ъю ъ тёяыхёъґ ґЁютэ ярёёшюэрЁэюёҐш, эю ш ╙ єЁюьюёюьх Ґюцх фюёҐр╕Ґё . ╥Ёґфэю ёърч𥹠яЁюшёєюфшҐ юфэр ьґҐрІш шыш эхёъюы№ъю ёЁрчґ, эю т яюёыхфґ■їшщ яхЁшюф ¤ъёярэёшш яЁюшёєюфшҐ ЁюёҐ ішёыхээюёҐш эютюую ёґяхЁ¤Ґэюёр ш ярфхэшх ішёыхээюёҐш ёґяхЁ¤Ґэюёют яЁхф√фґїхщ ¤яюєш. ┼фшэёҐтхээющ Іхээющ фюс√іхщ т Ґх Ёрээшх тЁхьхэр с√ыш цхэїшэ√ ш тё ъшщ Ёрч тхышъшщ тюцф№ яыхьхэш юёҐрты ы чр ёюсющ яЁртю юсїхэш ё яЁхъЁрёэющ яюыютшэющ яыхьхэш. |  |
  
|
|
╬ҐтхҐют - 47
, ёҐЁ:
1
2
3
All
[Ґюы№ъю эют√х]
|
|
|
|
| |
╧юёҐ N: 170
╟рЁхушёҐЁшЁютрэ: 24.02.13
╟рьхірэш :
|
|
╬ҐяЁртыхэю: 08.10.13 18:53. ╟руюыютюъ: ╤ тючэшъэютхэшхь яЁш..
╤ тючэшъэютхэшхь яЁшётрштр■їшє ЄюЁь єюч щёҐт𠤥эшіхёъшх яЁюІхёё√ Ёхчъю шчьхэшышё№. ╬єюҐэшъш ш ёюсшЁрҐхыш, яЁш тёхщ шє ҐюуфрЇэхщ ьэюуюішёыхээюёҐш ш ЇшЁюірщЇхь ЁрёяЁюёҐЁрэхэшш, ёҐрыш ёшёҐхьрҐшіхёъш ґёҐґя𥹠чхьыхфхы№Ірь ш ёъюҐютюфрь. ╚ юя Ґ№ тё╕ с√ыю чрт чрэю э𠤥эюухэхҐшіхёъшх яЁюІхёё√ ёґяхЁ¤Ґэюёют, схч ъюҐюЁ√є эх тючьюцэ√ ъръшх ышсю эютрІшш ъръ т ьрҐхЁшры№эющ, Ґръ ш т фґєютэющ юсырёҐш. ┬ёх фюёҐшцхэш іхыютхіхёҐтр яЁюшёєюфшыш т ръҐштэющ Єрчх ¤Ґэюухэхчр, ірёҐшіэю т шэхЁІшюээющ. ┬ уюьхюёҐрчх схчЁрчфхы№эю уюёяюфёҐтґхҐ ҐЁрфшІш .
┬хЁю ҐэюёҐ№ ьґҐрІшщ, т Ґюь ішёых ш т Y єЁюьюёюьх ьюцхҐ яЁюшчющҐш т ы■сюх тЁхь , тюҐ Ґюы№ъю т шэъґсрІшюээ√щ яхЁшюф ¤Ґр тхЁю ҐэюёҐ№ Ёхчъю тючЁрёҐрхҐ.
┬ шэхЁІшюээ√щ яхЁшюф шыш т уюьхюёҐрчх Ґрър ьґҐрІш эх сґфхҐ шьхҐ№ ъръшє ышсю чэрішҐхы№э√є яюёыхфёҐтшщ. ┬ шэъґсрІшюээ√щ яхЁшюф шыш т яхЁшюф ¤ъёярэёшш эюёшҐхыш Ґръющ ьґҐрІшш сыруюфрЁ т√ёюъюьґ ґЁютэ■ ярёёшюэрЁэюёҐш яюыґір■Ґ яхЁхф ёюёхф ьш чэрішҐхы№э√х яЁхшьґїхёҐтр, іҐю яЁштюфшҐ ъ ЁюёҐґ ішёыхээюёҐш фрээющ уряыюуЁґяя√ яЁш ёэшцхэшш фюыш фЁґушє уряыюуЁґяя.
| |
|
|
|
| |
═х чрЁхушёҐЁшЁютрэ
╟рЁхушёҐЁшЁютрэ: 01.01.70
|
|
╬ҐяЁртыхэю: 10.10.13 09:15. ╟руюыютюъ: ╨хъґрщ яшЇхҐ: ┬ё╕ ¤..
╨хъґрщ яшЇхҐ: | ІшҐрҐр: | | ┬ё╕ ¤Ґю ёыхф√ фЁхтэшє яЁюІхёёют ¤Ґэюухэхчр. |
|
╩ёҐрҐш ёърчрҐ№, тюҐ ¤Ґю тхё№ьр яхЁёяхъҐштэр ь√ёы№. ┼хх ёҐюшҐ яюфґь𥹠сюыхх фхҐры№эю. ╨хъґрщ яшЇхҐ: | ІшҐрҐр: | ┼фшэёҐтхээющ Іхээющ фюс√іхщ т Ґх Ёрээшх тЁхьхэр с√ыш цхэїшэ√ ш тё ъшщ Ёрч тхышъшщ тюцф№ яыхьхэш юёҐрты ы чр ёюсющ яЁртю юсїхэш ё яЁхъЁрёэющ яюыютшэющ яыхьхэш.
|
|
═ґ ґц ш хфшэёҐтхээющ?  ╥хь сюыхх, іҐю яЁш ръҐштэюь шэъюЁяюЁшЁютрэшш іЁхчт√ірщэю тхышъ Ёшёъ єшьхЁшчрІшш, ҐґЁъш тюҐ э𠤥юь ёярышышё№... | |
|
|
|
| |
╧юёҐ N: 632
╟рЁхушёҐЁшЁютрэ: 24.02.13
╟рьхірэш :
|
|
╬ҐяЁртыхэю: 27.04.15 19:38. ╟руюыютюъ: ┬ іхёырт яшЇхҐ: ═ґ ..
┬ іхёырт яшЇхҐ: ╫Ґю ьюцэю тч Ґ№ ё яыхьхэш, цштґїхую Ґхь, іҐю чр фхэ№ ёюсхЁ╕Ґ? └ шьхээю юс ¤Ґшє Ёрээшє ёҐрфш є юєюҐ√ ш ёюсшЁрҐхы№ёҐтр шф╕Ґ Ёхі№. ╠рҐхЁшры№эр ъґы№ҐґЁр яЁръҐшіхёъш юҐёґҐёҐтґхҐ. ╨хъюьхэфґ■ яюішҐрҐ№ л╧хіры№э√х ҐЁюяшъш╗ ╦хтш ╤ҐЁюёёр | |
|
|
|
| |
═х чрЁхушёҐЁшЁютрэ
╟рЁхушёҐЁшЁютрэ: 01.01.70
|
|
╬ҐяЁртыхэю: 28.04.15 12:31. ╟руюыютюъ: ╨хъґрщ яшЇхҐ: ╫Ґю ь..
╨хъґрщ яшЇхҐ: | ІшҐрҐр: | | ╫Ґю ьюцэю тч Ґ№ ё яыхьхэш, цштґїхую Ґхь, іҐю чр фхэ№ ёюсхЁ╕Ґ? |
|
╠эюую іхую. ╥хь сюыхх, іҐю хёыш "іҐю чр фхэ№", Ґю ¤Ґю эх яыхь фрцх, р ёъюЁхх ҐЁшср яЁшьрҐют. ┬ёх, іҐю т√Їх, ґцх шьххҐ эхъшх фюяюыэшҐхы№э√х ЁхёґЁё√, яЁхфёҐрты ■їшх шэҐхЁхё (ъЁюьх ҐЁґфют√є ш Ґюую ь ёр, іҐю эр эшє ёрьшє  ). | |
|
|
|
| |
╧юёҐ N: 172
╟рЁхушёҐЁшЁютрэ: 24.02.13
╟рьхірэш :
|
|
╬ҐяЁртыхэю: 10.10.13 18:25. ╟руюыютюъ: ърЁшс√ юіхэ№ фрцх рЁ..
ърЁшс√ юіхэ№ фрцх рЁртръёъшє цхэїшэ т ышіэ√є Іхы є шёяюы№чютрыш эр └эҐшырє, фрцх цхэёъшщ ч√ъ с√ы рЁртръёъшщ.
| |
|
|
|
| |
╧юёҐ N: 174
╟рЁхушёҐЁшЁютрэ: 24.02.13
╟рьхірэш :
|
|
╬ҐяЁртыхэю: 11.10.13 22:40. ╟руюыютюъ: ┼ёыш яЁхфяюыюцшҐ№, і..
┼ёыш яЁхфяюыюцшҐ№, іҐю эхсюы№Їр уЁґяяр эюёшҐхыхщ уряыюуЁґяя√ ┬ яюярыр т └чш■, р Ґюіэхх т └Ёртш■ ш уфх Ґю Ґрь яЁшъы■ішырё№ ьшъЁюърҐрёҐЁюЄр, Ґю юёэютрҐхы№ эютюую ёґяхЁ¤Ґэюёр ёєыюяюҐры эх Ґюы№ъю т√ёюъшщ ґЁютхэ№ ярёёшюэрЁэюёҐш, эю ш ьґҐрІшш M168 тьхёҐх ё P9.1 ш M294. ╧юёыхфґ■їшщ яхЁшюф ¤ъёярэёшш яЁштюфшҐ ъ Ґюьґ, іҐю ішёыхээюёҐ№ эрёхыхэш ёэшцрхҐё . ╧Ёш ¤Ґюь фюы эютюую ёґяхЁ¤Ґэюёр тючЁрёҐрхҐ чрёі╕Ґ ёэшцхэш ішёыхээюёҐш юъЁґцр■їхую эрёхыхэш . ╩ырёёшіхёъшщ ¤ЄЄхъҐ сґҐ√ыюіэюую уюЁы√Їър, ъюҐюЁ√щ эх яхЁхцшы ЁюфшҐхы№ёъшщ уряыюҐшя ┬, р эюёшҐхыш уряыюҐшяр ╤ юҐяЁртшышё№ эр тюёҐюъ чрёхы Ґ№ яЁшсЁхцэ√х Ёрщюэ√ └чшш ш └тёҐЁрышш. ╧ютҐюЁхэшх ьшъЁюърҐрёҐЁюЄ√ яЁштхыю ъ Ґюьґ, іҐю юёэютрҐхы№ эютюую ёґяхЁ¤Ґэюёр ёєыюяюҐры эх Ґюы№ъю т√ёюъшщ ґЁютхэ№ ярёёшюэрЁэюёҐш, эю ш ьґҐрІшш | ІшҐрҐр: |
├ряыюуЁґяяр D (M174) Ч уряыюуЁґяяр ─═╩ Y-єЁюьюёюь√. ╩ръ уряыюуЁґяяр D, Ґръ ш E ёюфхЁцрҐ юфэюэґъыхюҐшфэ√щ яюышьюЁЄшчь M168, ъюҐюЁ√щ шьххҐё тю тёхє уряыюуЁґяярє, ъЁюьх A ш B (ёь. уЁґяяр CT), р Ґръцх яюышьюЁЄшчь ґэшъры№эюую ёюс√Ґш YAP, ґэшъры№э√щ фы уряыюуЁґяя√ DE Ч юсїхую яЁхфър D ш ┼.
├ряыюуЁґяяр DE Ч ¤Ґю іхыютхіхёър Y-єЁюьюёюьэр уряыюуЁґяяр. ┼╕ юяЁхфхы ■Ґ ьґҐрІшш юфэюэґъыхюҐшфэюую яюышьюЁЄшчьр M1(YAP), M145(P205), M203, P144, P153, P165, P167, P183. |
|
╟р юфшэ Ёрч юэ шє ёєыюяюҐры шыш эхҐ, ёърч𥹠ҐЁґфэю. ╥юы№ъю ЁюфшҐхы№ёър уряыюуЁґяяр ╤ ґёяхыр ЇшЁюъю ЁрёяЁюёҐЁрэшҐ№ё ш сыруюяюыґіэю яхЁхцшыр ¤ъёярэёш■ ёґяхЁ¤Ґэюёр эюёшҐхы уряыюуЁґяя√ DE | |
|
|
|
| |
╧юёҐ N: 175
╟рЁхушёҐЁшЁютрэ: 24.02.13
╟рьхірэш :
|
|
╬ҐяЁртыхэю: 11.10.13 23:19. ╟руюыютюъ: ┬ючтЁрїхэшх сыґфэюую..
┬ючтЁрїхэшх сыґфэюую ё√эр.
╧ЁхфюёҐртшь эюёшҐхыхщ "уряыюҐшяр ╤" шє ёґф№сх т яюёыхфґ■їшь эхы╕уъюь ш чряґҐрээюь фхых тхҐтыхэш .
╧юяЁюсґхь ЁхъюэёҐЁґшЁют𥹠ёюс√Ґш ёт чрээ√х ё эюёшҐхы ьш уряыюҐшяр D.
╤ъюЁхх тёхую ірёҐ№ шє тхЁэґырё№ т └ЄЁшъґ, Ґръ ёърч𥹠эр "╚ёҐюЁшіхёъґ■ ╨юфшэґ". ╤ІхэрЁшщ ЁрчтшҐш ёюс√Ґшщ ёҐрэфрЁҐэ√щ. ╠шъЁюърҐрёҐЁюЄр, сґҐ√ыюіэюх уюЁы√Їъю, шёіхчэютхэшх рЄЁшърэёъюую Єшышрыр уряыюҐшяр D т яхЁшюф ¤ъёярэёшш ёґяхЁ¤Ґэюёр эюёшҐхы уряыюҐшяр ┼, яюёыхфґ■їшщ яхЁхєюф эр яЁюшчтюф їшх ЄюЁь√ єюч щёҐтютрэш , тҐюЁшёэ√щ юіру ¤Ґэюухэхчр ёЁхфш яЁхфёҐртшҐхыхщ эхуЁюшфэющ Ёрё√ ё чръюэюьхЁэ√ь т√Ґхёэхэшхь уряыюҐшяют └ ш ┬ ш яютёхьхёҐэ√ь ЁрёяЁюёҐЁрэхэшхь уряыюҐшяр ┼.
╤ІхэрЁшш ЁрчтшҐш ёюс√Ґшщ, яюёҐЁюхээ√х эр ҐхюЁшш ярёёшюэрЁэюёҐш, шёъы■ір■Ґ трЁшрэҐ ё тючэшъэютхэшхь ьґҐрІшш Y-─═╩ уфх Ґю Ґрь э𠸥юЁюэх ё яюёыхфґ■їшь ьрёёют√ь яхЁхёхыхэшхь эр эютюх ьхёҐю цшҐхы№ёҐтр. ╩ышьрҐшіхёъшх шчьхэхэш , яЁштюф їшх ъ ьрёёют√ь яхЁхёхыхэш ь эрЁюфют юс√іэю эх яЁштюф Ґ ъ ьшъЁюърҐръышчьрь, тёяыхёърь ярёёшюэрЁэюёҐш ш ьрёёют√ь ьґҐрІш ь, шфґҐ яЁюІхёё√ яЁшҐюър шыш юҐҐюър эрёхыхэш эр фрээющ ҐхЁЁшҐюЁшш. ╟рҐю т√ёюър тхЁю ҐэюёҐ№ шёіхчэютхэш яЁхфъют√є уряыюҐшяют эр ьхёҐх тючэшъэютхэш ьшъЁюърҐрёҐЁюЄ ё яюёыхфґ■їшьш тёяыхёърьш ярёёшюэрЁэюёҐш ш ьрёёют√ьш ьґҐрІш ьш эхяюёЁхфёҐтхээю т чюэх яюёыхфґ■їхщ ¤ъёярэёшш ёґяхЁ¤Ґэюёр, эюёшҐхы эютющ ьґҐрІшш Y-─═╩.
| |
|
|
|
| |
╧юёҐ N: 215
╟рЁхушёҐЁшЁютрэ: 24.02.13
╟рьхірэш :
|
|
╬ҐяЁртыхэю: 02.11.13 23:56. ╟руюыютюъ: ▌ЄЄхъҐ сґҐ√ыюіэюую у..
▌ЄЄхъҐ сґҐ√ыюіэюую уюЁы√Їър т яырэх Ёхчъюую ёэшцхэш ішёыхээюёҐш эрёхыхэш ьюу ґёшышт𥹸 чр ёі╕Ґ тючэшъэютхэш эют√є ЇҐрььют ёҐрЁ√є сюыхчэхщ, юҐ ъюҐюЁ√є эх с√ыю шььґэшҐхҐр.
╬ҐэюЇхэшх сръҐхЁшщ ш шє эюёшҐхыхщ яюєюцх эр юҐэюЇхэш ярЁрчшҐют ш шє єюч хт. ┴ґЁэюх ЁрёяЁюёҐЁрэхэшх тхф╕Ґ ъ т√ьшЁрэш■ ъюЁьютющ срч√, єюЁюЇшщ шььґэшҐхҐ уЁючшҐ ушсхы№■ сръҐхЁш ь.
┬ яхЁшюф ьюїэюую тёяыхёър шчыґіхэшщ т ¤яшІхэҐЁх ¤Ґэюухэхчр яхЁт√ьш ьґҐшЁґ■Ґ ьшъЁююЁурэшчь√, чрҐхь яЁюшёєюф Ґ ьшъЁюьґҐрІшш т юЁурэшчьрє ы■фхщ, ґёяхтЇшє т√ЁрсюҐрҐ№ шььґэшҐхҐ. ─юёҐр╕Ґё ш Y єЁюьюёюьх. ┬ Ёхчґы№ҐрҐх эюёшҐхы№ эютюую уряыюҐшяр яюыґірхҐ чэрішҐхы№э√х яЁхшьґїхёҐтр т ьрыюішёыхээющ юсїшэх юєюҐэшъют ш ёюсшЁрҐхыхщ. ┬ ёыґір є ё т√ёюъющ яыюҐэюёҐ№■ ш ьэюуюішёыхээюёҐ№■ эрёхыхэш Ґръшх яЁхшьґїхёҐтр яюыґірхҐ "сґъхҐ" уряыюҐшяют, юърчртЇшєё т ¤яшІхэҐЁх ¤Ґэюухэхчр.
| |
|
|
|
| |
╧юёҐ N: 467
╟рЁхушёҐЁшЁютрэ: 24.02.13
╟рьхірэш :
|
|
╬ҐяЁртыхэю: 28.08.14 13:25. ╟руюыютюъ: http://www.yfull.com..
| |
|
|
|
| |
╧юёҐ N: 472
╟рЁхушёҐЁшЁютрэ: 24.02.13
╟рьхірэш :
|
|
╬ҐяЁртыхэю: 08.09.14 00:49. ╟руюыютюъ: ═х т√єюфшы іхыютхъ ш..
| |
|
|
|
| |
╧юёҐ N: 473
╟рЁхушёҐЁшЁютрэ: 24.02.13
╟рьхірэш :
|
|
╬ҐяЁртыхэю: 08.09.14 16:27. ╟руюыютюъ: ╧ю ьэхэш■ ╩ы╕ёютр яЁ..
╧ю ьэхэш■ ╩ы╕ёютр яЁхфъш іхыютхър цшыш уфх Ґю Ґрь т ┼тЁрчшш.
╤эрірыр юҐ юсїхую ёҐтюыр юҐяюіъютрышё№ └00, яЁюэшъыш т └ЄЁшъґ ш чфхё№ ёюєЁрэ ышё№ т Ґхіхэшх яюёыхфэшє 200 Ґ√ё і ыхҐ.
╟рҐхь юҐ юсїхую ёҐтюыр юҐфхышышё№ └0, юэш Ґръ цх яЁюэшъыш т └ЄЁшъґ ыхҐ ¤фръ 180 Ґ√ё і эрчрф.
╧юҐюь т └ЄЁшъх юёхыш └1р, ёыхфюь └1b1
╬ Ґюь, іҐю т └ЄЁшъґ чрҐхь яЁюэшъыш ┬, ш чртхЁЇшыюё№ ¤Ґю яЁюэшъэютхэшхь ┼, └эрҐюышщ ґцх эх яшЇхҐ.
╧Ёш і╕ь эшъръшє ёыхфют юҐ яЁхс√трэш юсїхую ёҐтюыр уфх Ґю Ґрь т ┼тЁрчшш яюіхьґ Ґю эх ёюєЁрэшыюё№.
╚Ґюую ЇхёҐ№ ёыґірхт ґєюфр т └ЄЁшъґ яЁш яюыэюь юҐёґҐёҐтшш ёыхфют юҐ шє яЁшс√трэш . ╩ръ Ґю Ґръ т√єюфшҐ.
| |
|
|
|
|
|
| |
╧юёҐ N: 516
╟рЁхушёҐЁшЁютрэ: 24.02.13
╟рьхірэш :
|
|
╬ҐяЁртыхэю: 28.11.14 22:37. ╟руюыютюъ: ╫Ґю хёыш CF ш D - ..
| ІшҐрҐр: | ╫Ґю хёыш CF ш D - Ёрчэ√х тюыэ√ т√єюфр шч └ЄЁшъш?
╫Ґю хёыш DE тючэшъыр т └ЄЁшъх?
╫Ґю хёыш E тючэшъыр т └ЄЁшъх?
╫Ґю хёыш Ґюы№ъю юҐфхы№э√х ёґсъырф√ E т√Їыш шч └ЄЁшъш т фЁхтэюёҐш юіхэ№ яючфэю (т эхюышҐх шыш эхёъюы№ъю Ёрэхх)? |
|
┬хЁю ҐэюёҐ№ ьґҐрІшщ, т Ґюь ішёых ш т Y єЁюьюёюьх ьюцхҐ яЁюшчющҐш т ы■сюх тЁхь , тюҐ Ґюы№ъю т шэъґсрІшюээ√щ яхЁшюф ¤Ґр тхЁю ҐэюёҐ№ Ёхчъю тючЁрёҐрхҐ. ╧Ёшішэр тю тэхЇэхь тючфхщёҐтшш эр яюяґы Іш■, чряґёър■їґ■ яЁюІхёё√ ¤Ґэюухэхчр. ┬ шэхЁІшюээ√щ яхЁшюф шыш т уюьхюёҐрчх Ґрър ьґҐрІш эх сґфхҐ шьхҐ№ ъръшє ышсю чэрішҐхы№э√є яюёыхфёҐтшщ. ┬ёх ьґцёъшх ышэшш шьх■Ґ сюыхх эх ьхэхх Ёртэ√х Їрэё√ эр ёґїхёҐтютрэшх. ┬ шэъґсрІшюээ√щ яхЁшюф ш т яхЁшюф ¤ъёярэёшш эюёшҐхыш Ґръющ ьґҐрІшш сыруюфрЁ т√ёюъюьґ ґЁютэ■ ярёёшюэрЁэюёҐш яюыґір■Ґ яхЁхф ёюёхф ьш чэрішҐхы№э√х яЁхшьґїхёҐтр, іҐю яЁштюфшҐ ъ ЁюёҐґ ішёыхээюёҐш фрээющ уряыюуЁґяя√ яЁш ёэшцхэшш фюыш фЁґушє уряыюуЁґяя. ╧юыґірхҐё Ёхчъюх ёюъЁрїхэшх, р яЁш эшчъющ ішёыхээюёҐш яюяґы Ішш ш яюыэюх шёіхчэютхэшх ёҐрЁ√є, эх ьґҐшЁютртЇшє, ышэшщ яЁш ръҐштэюь ЁюёҐх фюыш эютющ ышэшш, яюыґіштЇхщ т фютхёюъ юҐ юфэющ фю эхёъюы№ъшє хфшэютЁхьхээ√є ьґҐрІшщ т Y єЁюьюёюьх. ╧юяґы Іш яЁюєюфшҐ іхЁхч сґҐ√ыюіэюх уюЁы√Їъю, іҐю тхф╕Ґ ъ чэрішҐхы№эюьґ шчьхэхэш■ сґъхҐр Y єЁюьюёюь. ─юы ЁхышъҐют√є, эх ьґҐшЁютртЇшє, єЁюьюёюь Ёхчъю ёюъЁрїрхҐё чр ёі╕Ґ тючЁрёҐрэш ьґҐшЁютртЇшє. ╒юіґ юсЁрҐшҐ№ тэшьрэшх, іҐю Ґръшх ьґҐрІшш ты ■Ґё фюяюыэшҐхы№э√ь яЁшыюцхэшхь, яюсюіэ√ь ¤ЄЄхъҐюь, ъ т√ёюъюьґ ґЁютэ■ ярёёшюэрЁэюёҐш шє эюёшҐхыхщ. ╤рьш яю ёхсх юэш эш ъръющ Ёюыш эх шуЁр■Ґ. ┬хЁэ╕ьё т ъюэҐръҐэґ■ чюэґ └ЄЁшъш ш └чшш. ┼ёыш с√ ╤ тючэшъыр т └ЄЁшъх, Ґю т яхЁшюф ¤ъёярэёшш юэр с√ яюыґішыр эр ъюэҐшэхэҐх ЇшЁюъюх ЁрёяЁюёҐЁрэхэшх. ┬ яюёыхфґ■їхь тхЁю ҐэюёҐ№ х╕ яюыэюую шёіхчэютхэш т Ёхчґы№ҐрҐх яюёыхфґ■їшє ьшъЁюърҐрёҐЁюЄ єюҐ ш ьюуыр с√Ґ№ т√ёюъющ, эю уфх ышсю ьюуыр ш ёюєЁрэшҐ№ё . ╬фэръю Ґръшє ёыґірхт яюър эх юсэрЁґцхэю. ╬я Ґ№ цх яютҐюЁэюх яюыэюх шёіхчэютхэшх т └ЄЁшъх D Ґюцх тючьюцэю, эю хї╕ ьхэхх тхЁю Ґэю. ╧Ёхфяюыюцшь, іҐю т └чш■ яЁюэшъыр эхсюы№Їр яюяґы Іш ┬ ш эх ґёяхыр ЇшЁюъю ЁрёяЁюёҐЁрэшҐ№ё яю ьрҐхЁшъґ, Ґюіэхх яю └Ёртшш. ╧ЁшсЁхцэ√х ёюсшЁрҐхыш ьюуыш с√Ґ№ срэры№эю яЁшцрҐ√ яґёҐ√эхщ ъ схЁхуґ. ╩ Ґюьґ цх т шэхЁІшюээ√щ яхЁшюф яюфтшцэюёҐ№ эрёхыхэш эшчър . ╧хЁтюх цх яЁюєюцфхэшх іхЁхч сґҐ√ыюіэюх уюЁы√Їъю ьюуыю яЁштхёҐш ъ яюыэюьґ шёіхчэютхэш■ ЁхышъҐютюую ┬, чрьхэющ хую эр ╤. ╧хЁшюф ¤ъёярэёшш яюфЁрчґьхтрхҐ т√ёюъґ■ яюфтшцэюёҐ№ эрёхыхэш . ┬ яюёыхфґ■їшх ¤яюєш ╤ ръҐштэю ЁрёяЁюёҐЁрэ хҐё яю ьрҐхЁшъґ. ╤шёҐхьрҐшіхёъшх ьшъЁюърҐрёҐЁюЄ√ ёяюёюсёҐтютрыш ъръ фры№эхщЇхьґ Ёрёёхыхэш■, Ґръ ш ґёяхЇэюьґ тхҐтыхэш■.
| |
|
|
|
| |
╧юёҐ N: 630
╟рЁхушёҐЁшЁютрэ: 24.02.13
╟рьхірэш :
|
|
╬ҐяЁртыхэю: 27.04.15 02:11. ╟руюыютюъ: ґ сЁрьшэют ╟рярфэющ ..
ґ сЁрьшэют ╟рярфэющ ┴хэурышш фюы уряыюуЁґяя√ R1a Ц 72 %, ґ сЁрьшэют ЇҐрҐр ╙ҐҐрЁ ╧ЁрфхЇ Ц 68 %
| |
|
|
|
| |
═х чрЁхушёҐЁшЁютрэ
╟рЁхушёҐЁшЁютрэ: 01.01.70
|
|
╬ҐяЁртыхэю: 27.04.15 09:59. ╟руюыютюъ: ╨хъґрщ яшЇхҐ: ґ сЁр..
╨хъґрщ яшЇхҐ: | ІшҐрҐр: | | ґ сЁрьшэют ╟рярфэющ ┴хэурышш фюы уряыюуЁґяя√ R1a Ц 72 %, ґ сЁрьшэют ЇҐрҐр ╙ҐҐрЁ ╧ЁрфхЇ Ц 68 % |
|
└ тюҐ шэҐхЁхёэ√щ ьюьхэҐ: юфшэ шч ъы■іхт√є Ґхчшёют шэфґшёҐёъющ ЁхЄюЁьрІшш с√ы ъръ Ёрч т юҐърчх сЁрьшэрь т шє эрёыхфёҐтхээ√є яЁртрє. ╟рэ Ґэю, эх Ґръ ыш? | |
|
|
|
| |
╧юёҐ N: 633
╟рЁхушёҐЁшЁютрэ: 24.02.13
╟рьхірэш :
|
|
╬ҐяЁртыхэю: 28.04.15 21:20. ╟руюыютюъ: ╧ЁръҐшіхёъш тёх уряы..
└ ╩ы╕ёют | ІшҐрҐр: | | ╧ЁръҐшіхёъш тёх уряыюуЁґяя√-Ёюфр шч ┼тЁюя√ яЁюярыш. ─═╩-рЁєхюыюуш тшфшҐ Ёхчъґ■ ёьхэґ эрёхыхэш ┼тЁюя√ т III Ґ√ё. фю э. ¤. ▌Ґю цх тшфшҐ ─═╩-ухэхрыюуш , яю фрээ√ь ъюҐюЁющ э√эхЇэхх эрёхыхэшх ┼тЁюя√ люсэґы хҐё ╗ яЁшьхЁэю 4500 ыхҐ эрчрф, ш яЁюєюфшҐ, ъръ уютюЁ Ґ ухэхҐшъш, лсґҐ√ыюіэюх уюЁы√Їъю яюяґы Ішш╗. ╚ёіхчыр уряыюуЁґяяр G2a, ёрьр ЁрёяЁюёҐЁрэхээр т л╤ҐрЁющ ┼тЁюях╗, ш уфх эш ъюярыш ─═╩-рЁєхюыюуш, юэр тхчфх яЁю ты ырё№ фю 5000 ыхҐ эрчрф. ╧юҐюь юэр тфЁґу шч ┼тЁюя√ шёіхчыр ш юс· тшырё№ т ╠хёюяюҐрьшш, эр ╩ртърчх, т ╧хЁхфэхщ └чшш, ёю тЁхьхэрьш юсїхую яЁхфър яЁшьхЁэю 4500Ц4000 ыхҐ эрчрф, Ґю хёҐ№ яЁющф Ґю ёрьюх лсґҐ√ыюіэюх уюЁы√Їъю яюяґы Ішш╗ ш эрірт тючЁюцфхэшх яЁръҐшіхёъш ё эґы . ▌Ґю, хёыш ґуюфэю, ¤ЄЄхъҐ ляюёыхфэхую шч ьюушърэ╗, ъюуфр яюёыхфэшщ т√цшы ш фры т√цштЇхх яюҐюьёҐтю, ёҐрт Ґръшь юсЁрчюь эют√ь люсїшь яЁхфъюь╗ яюяґы Ішш. ╧Ёюярыр уряыюуЁґяяр E-V13, ъюҐюЁр яю Ёрёъюяърь цшыр т ┼тЁюях 7 Ґ√ё і ыхҐ эрчрф. ═√эхЇэхх эрёхыхэшх E-V13 т ┼тЁюях (ш тхчфх) эрішэрхҐё юҐ юсїхую яЁхфър, цштЇхую 3600 ыхҐ эрчрф. ╧Ёюярыр уряыюуЁґяяр I1, ґ ъюҐюЁющ с√ыр фЁхтэхщЇр шёҐюЁш т ┼тЁюях, ш юсїшщ яЁхфюъ э√эхЇэшє эюёшҐхыхщ I1 яю тёхщ ┼тЁюях, юҐ └ҐырэҐшъш фю ╙Ёрыр, с√ы юфшэ ш ҐюҐ цх, ш цшы тёхую 3400 ыхҐ эрчрф. ╧Ёюярыш шч ┼тЁюя√ уряыюуЁґяя√ R1a ш I2a, эршсюыхх яЁхфёҐртшҐхы№э√х ёхщірё эр ┴рыърэрє ш т ╤хЁсшш, ш ь√ ёхщірё юёҐрэютшьё эр шє ёґф№сх сюыхх яюфЁюсэю. |
|
| ІшҐрҐр: | | ├ряыюуЁґяяр I2a ЁрчюЁтрырё№ яюяюырь Ц юфэр ірёҐ№ схцрыр эр ┴ЁшҐрэёъшх юёҐЁютр, фЁґур Ц эр ─ґэрщ. ╧юэрфюсшыюё№ сюы№Їх фтґє Ґ√ё і ыхҐ, ё 4500 фю 2300 ыхҐ эрчрф, іҐюс√ уряыюуЁґяяр I2a т фґэрщёъюь Ёхушюэх яюЇыр т ЁюёҐ, ш ёҐрыр чрёхы Ґ№ ┬юёҐюіэґ■ ┼тЁюяґ. ─═╩-ухэхрыюушіхёъшх ЁрёіхҐ√ яюърч√тр■Ґ, іҐю тёх I2a тю тёхщ ┬юёҐюіэющ ┼тЁюях Ц юҐ ├ЁхІшш эр ■ух фю ╧ЁшсрыҐшъш эр ёхтхЁх, шьх■Ґ юфэюую юсїхую яЁхфър, ъюҐюЁ√щ цшы 2300 ыхҐ эрчрф, т ъюэІх яЁюЇыющ ¤Ё√, тшфшью, эр ─ґэрх. |
|
| |
|
|
|
| |
╧юёҐ N: 643
╟рЁхушёҐЁшЁютрэ: 24.02.13
╟рьхірэш :
|
|
╬ҐяЁртыхэю: 23.05.15 20:41. ╟руюыютюъ: "╧Ёш сюыхх сышчъ..
"╧Ёш сюыхх сышчъюь ЁрёёьюҐЁхэшш ётхфхэшщ юс шёъюярхь√є уряыюҐшярє т√Ёшёют√трхҐё ърЁҐшэр яюіҐш яюыэюую шёіхчэютхэш ьґцёъшє уряыюуЁґяя т ╤ҐрЁющ ┼тЁюях т Ґхіхэшх III Ґ√ё. фю э.¤., яю ёҐЁрээюьґ ёютярфхэш■ т єюфх чрёхыхэш ╓хэҐЁры№эющ ш ╟рярфэющ ┼тЁюя√ ¤Ёсшэрьш, эюёшҐхы ьш уряыюуЁґяя√ R1b. ╧Ёюярыш уряыюуЁґяя√ G2a, E1b-V13, I1, I2, R1a, ш ю эшє ь√ чэрхь Ґюы№ъю яюҐюьґ, іҐю юэш яЁюярыш эх яюыэюёҐ№■, р юёҐрҐъш шє схцрыш шч ╓хэҐЁры№эющ ┼тЁюя√ т Ёрчэ√х ъЁр . R1a схцрыш эр ╨ґёёъґ■ Ёртэшэґ (ё юсїшь яЁхфъюь 4900 ыхҐ эрчрф), I2 Ц эр ┴ЁшҐрэёъшх юёҐЁютр (ё юсїшь яЁхфъюь юъюыю 5000 ыхҐ эрчрф) ш т ┬юёҐюіэґ■ ┼тЁюяґ (т√Їыш шч сґҐ√ыюіэюую уюЁы√Їър яюяґы Ішш Ґюы№ъю 2300 ыхҐ эрчрф, т ъюэІх яЁюЇыющ ¤Ё√). I1 яЁюярыр яюіҐш яюыэюёҐ№■, ш ёҐрыр тючЁюцф𥹸 Ґюы№ъю 3600 ыхҐ эрчрф. G2a схцрыш т ╠рыґ■ └чш■, ш юҐҐґфр ЁрчюЇышё№ яю ╠хёюяюҐрьшш ш ╧хЁхфэхщ └чшш, ш фтшэґышё№ эр ╩ртърч. ┬ шҐюух ёхщірё т ┼тЁюях фюы эюёшҐхыхщ G2a ёюёҐрты хҐ тёхую хфшэшІ√ яЁюІхэҐют, ш ¤Ґю тъы■ір ьэюушє тхЁэґтЇшєё т ┼тЁюяґ шє яюҐюьъют. ╥ръ іҐю эшъръющ лярыхюышҐшіхёъющ эхяЁхЁ√тэюёҐш╗ т ┼тЁюях эх с√ыю. ┴√ыр яюіҐш яюыэр чрьхэр ьґцёъшє уряыюуЁґяя ╤ҐрЁющ ┼тЁюя√ эр уряыюуЁґяяґ R1b ьхцфґ 4800 ш 4000 ыхҐ эрчрф, ш Ґюы№ъю т I Ґ√ё. фю э.¤. эюёшҐхыш фЁґушє уряыюуЁґяя ёҐрыш тючтЁрїрҐ№ё т ┼тЁюяґ." ┬ уряыюҐшях т√Їх юҐьхіхэ√ эршсюыхх яЁшэІшяшры№э√х Ёрёєюцфхэш т тхышішэрє рыыхыхщ яЁхфъют√є уряыюҐшяют ёґсъырфют G2a1a ш G2a3b, шє чфхё№ эрёішҐ√трхҐё 20 ьґҐрІшщ. ▌Ґю ґцх яюьхїрхҐ юсїхую яЁхфър фтґє ёґсъырфют эр 7000 ыхҐ эрчрф, ш ¤Ґю тЁхь цшчэш Ґюы№ъю юсїхую яЁхфър уЁґяя√ G2a эр ╩ртърчх. ┼їх уыґсцх яю тЁхьхэш, юъюыю 20 Ґ√ё і ыхҐ эрчрф, цшы юсїшщ яЁхфюъ G2, ш хїх фЁхтэхх, юъюыю 50 Ґ√ё і ыхҐ эрчрф Ц юсїшщ яЁхфюъ G. ╤ґсъырф G2a тяюёыхфёҐтшш ЁрчюЇхыё эр сюыхх ьюыюф√х тхҐтш, эю юэш эх Ёрчфхы ■Ґё яю яхЁхішёыхээ√ь ¤Ґэюёрь, тёх юёҐр■Ґё сЁрҐ№ ьш яю Ёюфґ. ╦шЇ№ т юфэющ тхҐтш, тючЁрёҐюь тёхую 450▒175 ыхҐ, эрєюф Ґё Ґюы№ъю юёхҐшэ√ Ц фшуюЁІ√ ш шЁюэІ√. ▌Ґю Ц фюіхЁэ тхҐт№ юёэютэющ, фюьшэшЁґ■їхщ тхҐтш, т ъюҐюЁющ 85% юҐ тёхє уряыюҐшяют уряыюуЁґяя√ G2a. ┬ючЁрёҐ ¤Ґющ фюьшэшЁґ■їхщ тхҐтш Ц 1675▒380 ыхҐ, Ґю хёҐ№ IV тхъ эрЇхщ ¤Ё√ яы■ё-ьшэґё іхҐ√Ёх тхър. ═р яхЁт√щ тчуы ф, ¤Ґю рырэёъшх тЁхьхэр. ═ю ЇҐґър т Ґюь, іҐю ¤Ґр фюьшэшЁґ■їр тхҐт№ яЁюшчюЇыр юҐ фЁхтэхщ тхҐтш ё тючЁрёҐюь 4-5 Ґ√ё і ыхҐ, ъюуфр эшъръшє рырэют эх с√ыю. ╙ ¤Ґющ фЁхтэхщ тхҐтш Ц ётю шёҐюЁш , хх уряыюуЁґяяр G2a тЁ ф ыш шьххҐ юҐэюЇхэшх ъ рырэрь-ёъшЄрь-ёрърь, уряыюуЁґяяр ъюҐюЁ√є с√ыр, ёъюЁхх тёхую, R1a. ╩ Ґюьґ цх ъюЁэш ¤Ґющ тхҐтш G2a яЁюёыхцштр■Ґё т ╚ёярэшш ш ╘ЁрэІшш, уфх шёъюярхь√х уряыюҐшя√ G2a, ъръ ґцх с√ыю ґяюь эґҐю, фрҐшЁґ■Ґё 7000 ыхҐ эрчрф. ═х ёішҐр ґяюь эґҐ√є юҐэюёшҐхы№эю ьюыюф√є тхҐтхщ, юсЁрчютртЇшєё т яхЁтюь Ґ√ё іхыхҐшш эрЇхщ ¤Ё√ шыш фрцх т ёЁхфэшх тхър, Іхы√щ Ё ф тхҐтхщ ёхтхЁю-чрярфэюую ш ІхэҐЁры№эюую ╩ртърчр схЁ╕Ґ ётюш шёҐюъш т эрірых, ёхЁхфшэх ш ъюэІх I Ґ√ё. фю эрЇхщ ¤Ё√. ▌Ґю Ц ґяюь эґҐ√х т√Їх ҐЁш тхҐтш уЁґчшэ, ё тючЁрёҐюь 2425▒270 ыхҐ эрчрф (ёґсъырф G2a1a1), 2175▒340 ыхҐ эрчрф (G2a1a1), 3025▒375 ыхҐ эрчрф (G2a3b1). ╧ЁшьхЁэю Ґръющ цх тючЁрёҐ шьх■Ґ тхҐтш іхЁъхёют, фшуюЁІхт, шЁюэІхт (2500▒490 ыхҐ), р Ґръцх рсєрчют, іхЁъхёют, фшуюЁІхт ш шЁюэІхт (2500▒430 ыхҐ фю юсїхую яЁхфър). ┴юы№ЇшэёҐтю ¤Ґшє Ёрёєюф їшєё тхҐтхщ яЁюфюыцр■Ґ Ёюф G2a-P18 эр ╩ртърчх, ъюҐюЁ√щ яю тшыё Ґрь ьхцфґ 4 ш 5 Ґ√ё ірьш ыхҐ эрчрф. ╥ю, іҐю ¤Ґш тхҐтш яЁюфюыцр■Ґё ґ юёхҐшэ ё Ґхє фртэшє тЁхьхэ, яюърч√трхҐ, іҐю эшъръшє яЁшЇхфЇшє ёю ёҐюЁюэ√ рырэют Ґрь эхҐ, тю тё ъюь ёыґірх, т чрьхҐэ√є ъюышіхёҐтрє. ▀ёэю ш Ґю, іҐю юёхҐшэ√ т юҐэюЇхэшш шє фюьшэшЁґ■їхщ уряыюуЁґяя√ эшіхь эх юҐышір■Ґё юҐ ётюшє ъртърчёъшє ёюёхфхщ. ═х ёішҐр ґяюь эґҐ√є юҐэюёшҐхы№эю ьюыюф√є тхҐтхщ, юсЁрчютртЇшєё т яхЁтюь Ґ√ё іхыхҐшш эрЇхщ ¤Ё√ шыш фрцх т ёЁхфэшх тхър, Іхы√щ Ё ф тхҐтхщ ёхтхЁю-чрярфэюую ш ІхэҐЁры№эюую ╩ртърчр схЁ╕Ґ ётюш шёҐюъш т эрірых, ёхЁхфшэх ш ъюэІх I Ґ√ё. фю эрЇхщ ¤Ё√. ▌Ґю Ц ґяюь эґҐ√х т√Їх ҐЁш тхҐтш уЁґчшэ, ё тючЁрёҐюь 2425▒270 ыхҐ эрчрф (ёґсъырф G2a1a1), 2175▒340 ыхҐ эрчрф (G2a1a1), 3025▒375 ыхҐ эрчрф (G2a3b1). ╧ЁшьхЁэю Ґръющ цх тючЁрёҐ шьх■Ґ тхҐтш іхЁъхёют, фшуюЁІхт, шЁюэІхт (2500▒490 ыхҐ), р Ґръцх рсєрчют, іхЁъхёют, фшуюЁІхт ш шЁюэІхт (2500▒430 ыхҐ фю юсїхую яЁхфър). ┴юы№ЇшэёҐтю ¤Ґшє Ёрёєюф їшєё тхҐтхщ яЁюфюыцр■Ґ Ёюф G2a-P18 эр ╩ртърчх, ъюҐюЁ√щ яю тшыё Ґрь ьхцфґ 4 ш 5 Ґ√ё ірьш ыхҐ эрчрф. ╥ю, іҐю ¤Ґш тхҐтш яЁюфюыцр■Ґё ґ юёхҐшэ ё Ґхє фртэшє тЁхьхэ, яюърч√трхҐ, іҐю эшъръшє яЁшЇхфЇшє ёю ёҐюЁюэ√ рырэют Ґрь эхҐ, тю тё ъюь ёыґірх, т чрьхҐэ√є ъюышіхёҐтрє. ▀ёэю ш Ґю, іҐю юёхҐшэ√ т юҐэюЇхэшш шє фюьшэшЁґ■їхщ уряыюуЁґяя√ эшіхь эх юҐышір■Ґё юҐ ётюшє ъртърчёъшє ёюёхфхщ.╨рёіхҐ яю 19-ьрЁъхЁэ√ь уряыюҐшярь фры 1675▒380 ыхҐ фю юсїхую яЁхфър тхҐтш, р яю 111-ьрЁъхЁэ√ь, яЁшіхь Ґюы№ъю фы юёхҐшэ Ц 1375▒210 ыхҐ, Ґю хёҐ№ Ґю цх ёрьюх т яЁхфхырє яюуЁхЇэюёҐш ЁрёіхҐют. ▌Ґю яЁшьхЁэю VII тхъ, яы■ё-ьшэґё ярЁр тхъют. ═ю ¤Ґю, яютҐюЁ хь, Ґюы№ъю юфэр, єюҐ№ ш юёэютэр тхҐт№, ш ъюҐюЁр юсЁрчґхҐ юсїґ■ ёшёҐхьґ тхҐтхщ ё юсїшь яЁхфъюь юъюыю 5 Ґ√ё і ыхҐ эрчрф. ┬ Іхыюь, юёхҐшэёъшх 67-ьрЁъхЁэ√х уряыюҐшя√ уЁґяя√ G2a1-P18 шьх■Ґ юсїхую яЁхфър эхёъюы№ъшє тхҐтхщ яЁшьхЁэю 3025 ыхҐ эрчрф. ▌Ґю Ц ъюэхІ II Ґ√ё іхыхҐш фю эрЇхщ ¤Ё√. ┬ ├Ёґчшш, ъръ юяшёрэю т√Їх Ц т уряыюуЁґяях G2a эр ҐЁш іхҐтхЁҐш уряыюҐшя√ Ґющ цх уЁґяя√ G2a1-╨18. ─юсртшь хїх эхёъюы№ъю ёыют ю ёґсъырфх G2a3b-P303 (ыхтр ірёҐ№ фхЁхтр эр Ёшё. т√Їх ш ыхтр тхЁєэ ірёҐ№ фхЁхтр ґ уЁґчшэёъшє уряыюҐшяют). G2a3b-P303 тъы■ірхҐ яЁръҐшіхёъш тёхє Їряёґуют, фтх ҐЁхҐш іхЁъхёют, 10% уЁґчшэ ш яюыютшэґ рсєрчют уряыюуЁґяя√ G2a. ┼ёыш яЁютхёҐш ЁрёіхҐ эх Ґюы№ъю яю уряыюҐшярь ├Ёґчшш, р яю тёхьґ ёхтхЁю-чрярфэюьґ ш ІхэҐЁры№эюьґ ╩ртърчґ, Ґю юёэютэ√х тхҐтш G2a1a1 ш G2a3b1 Ёрчфхы ■Ґё юъюыю 12 Ґ√ё і ыхҐ эрчрф. ┬юҐ Ґръ эрьэюую Ёрёєюф Ґё яю ётюхщ ─═╩-ухэхрыюушш юёхҐшэ√ юҐ рф√уют, Ґръ цх Ёрёєюф Ґё ш рсєрч√ тэґҐЁш ётюхую ¤Ґэюёр. ╧ю эхчртшёшь√ь ЁрёіхҐрь юсїшщ яЁхфюъ ёґсъырфр G2a цшы яЁшьхЁэю 12400 ыхҐ эрчрф (╩ы╕ёют, 2011). ╬я Ґ№, яюёъюы№ъґ ё Ґхє фртэшє яюЁ Ёюф G яЁюфюыцрхҐ ЁрёєюфшҐ№ё эр ёґсъырф√ ш тхҐтш тяыюҐ№ фю эрёҐю їхую тЁхьхэш, рырэрь Ґрь ьхёҐр эхҐ http://forumkavkaz.com/index.php?topic=9679.0 | |
|
|
|
| |
═х чрЁхушёҐЁшЁютрэ
╟рЁхушёҐЁшЁютрэ: 01.01.70
|
|
╬ҐяЁртыхэю: 03.06.15 13:02. ╟руюыютюъ: └ эх сґфхҐ ыш ┬рь шэ..
└ эх сґфхҐ ыш ┬рь шэҐхЁхёхэ т ¤Ґюь ъюэҐхъёҐх фшёёхЁ ├рыъшэющ ю ╨ґёёъюь ърурэрҐх? ╩эшцър хёҐ№ т сыюух ёрщҐр. ┬Ёюфх с√ ╩ыхёют эшіхую юс ¤Ґюь эх яшёры? ╚эҐхЁхёэю, іҐю т ърурэрҐх с√ыю яЁхфёҐртыхэю фтр іхҐъю фшЄЄхЁхэІшЁютрээ√є рэҐЁюяюыюушіхёъшє Ґшяр. └ фы ёхс эрЇхы, іҐю ьрҐхЁшры√ ьюушы№эшър ╩юЁхы√ шьх■Ґ тэ√х ярЁрыыхыш ё Ґшяшіэ√ьш чрєюЁюэхэш ьш тЁхьхэ ърурэрҐр.
| |
|
|
|
| |
═х чрЁхушёҐЁшЁютрэ
╟рЁхушёҐЁшЁютрэ: 01.01.70
|
|
╬ҐяЁртыхэю: 20.06.15 11:51. ╟руюыютюъ: ґ сЁрьшэют ╟рярфэющ ..
| ІшҐрҐр: | | ґ сЁрьшэют ╟рярфэющ ┴хэурышш фюы уряыюуЁґяя√ R1a Ц 72 %, ґ сЁрьшэют ЇҐрҐр ╙ҐҐрЁ ╧ЁрфхЇ Ц 68 % |
|
┬ююсїх эрфю ёьюҐЁхҐ№ шёҐюЁш■ ърёҐютющ ёҐрсшы№эюёҐш. ╫Ґю, ърёҐ√ эхшчьхээ√ ёю тЁхь╕э ╨шутхф√? └ тфЁґу ёютЁхьхээ√х ърёҐ√ Ёхчґы№ҐрҐ "яхЁхчруЁґчъш ёшёҐхь√" яюёых ьґёґы№ьрэёъшє тҐюЁцхэшщ, эряЁшьхЁ? ═рфю ёьюҐЁхҐ№ фюъґьхэҐры№э√х ухэхрыюушш... ╦хухэфрЁэ√х ухэхрыюушш тёхЁ№╕ч эх сЁрҐ№, шсю яЁютхЁшҐ№ ёыюцэю, Ґюы№ъю яюфышээ√х ухэхрыюушіхёъшх фюъґьхэҐ√ трцэ√ т яхЁтґ■ юіхЁхф№. ╥ю хёҐ№, т√ёюъшх яЁюІхэҐ√ R1a ё юфэющ ёҐюЁюэ√ тяхірҐы ■Ґ, эю эрфю ъюя𥹸 фры№Їх, т ёґсъыхщфрє, т фрҐшЁютърє, т ухэхрыюуш є... ╧хЁтюх тяхірҐыхэшх ьюцхҐ с√Ґ№ юсьрэішт√ь, юёюсхээю хёыш ъръшх-Ґю R1a тёҐЁхір■Ґё , хёыш ьэх эх шчьхэ хҐ ярь Ґ№, ґ ъръшє-Ґю яыхь╕э Ґшяр ёрєрЁшр (тЁюфх яю ч√ъґ ьґэфр) ш іхэіґ (фЁртшф√ яю ч√ъґ)... | |
|
|
|
| |
═х чрЁхушёҐЁшЁютрэ
╟рЁхушёҐЁшЁютрэ: 01.01.70
|
|
╬ҐяЁртыхэю: 20.06.15 11:58. ╟руюыютюъ: ┴рҐръ яшЇхҐ: юёюсхэ..
┴рҐръ яшЇхҐ: | ІшҐрҐр: | юёюсхээю хёыш ъръшх-Ґю R1a тёҐЁхір■Ґё , хёыш ьэх эх шчьхэ хҐ ярь Ґ№,
ґ ъръшє-Ґю яыхь╕э Ґшяр ёрєрЁшр (тЁюфх яю ч√ъґ ьґэфр) ш іхэіґ (фЁртшф√ яю ч√ъґ)... |
|
╚ьх■ ттшфґ, іҐю т рЁшхт-ьхурюёхьхэшҐхыхщ эх юіхэ№ тхЁ■. ╟ріхь с√ тхфшщёъшь сЁрєьрэрь т ьрёёх чрярф𥹠эр яюыґфшъшє Ґхьэюъюцшє рсюЁшухэюъ? └ тфЁґу ъръшх-Ґю R1a яЁюэшърыш т ╚эфш■ чрфюыую фю рЁшхт? └ тфЁґу сЁрєьрэ√ т юёэютэюь рсюЁшухээюую (эхрЁшщёъюую) яЁюшёєюцфхэш ? ╨рчсшЁрҐ№ё эрфю.  | |
|
|
|
| |
╧юёҐ N: 647
╟рЁхушёҐЁшЁютрэ: 24.02.13
╟рьхірэш :
|
|
╬ҐяЁртыхэю: 20.06.15 15:14. ╟руюыютюъ: ╚эфш ¤Ґю Ґръющ юуЁю..
╚эфш ¤Ґю Ґръющ юуЁюьэ√щ Ґґяшъ, ёґсъюэҐшэхэҐ, юъЁґц╕ээ√щ уюЁрьш. ╦■фёъшх яюҐюъш шчфЁхтых яЁюэшърыш Ґґфр, юсцштрышё№, юърч√трыш ётю╕ тыш эшх, шёя√Ґ√трыш тючфхщёҐтш эр ёхсх, юфэръю ьрыю ъҐю яюъшфры ¤ҐюҐ "юрчшё".
┼ї╕ эрфю шьхҐ№ т тшфґ, іҐю фышҐхы№эюх тЁхь ёґсъюэҐшэхэҐ шьхы ьэюую ьрыюэрёхы╕ээ√є ш яґёҐ√ээ√є ьхёҐ, юъЁґцртЇшє Ёхфъшх юіруш ІштшышчрІшш.
| |
|
|
|
| |
╧юёҐ N: 648
╟рЁхушёҐЁшЁютрэ: 24.02.13
╟рьхірэш :
|
|
╬ҐяЁртыхэю: 20.06.15 22:23. ╟руюыютюъ: ┼ V-13..
| |
|
|
|
|
|
| |
═х чрЁхушёҐЁшЁютрэ
╟рЁхушёҐЁшЁютрэ: 01.01.70
|
|
╬ҐяЁртыхэю: 20.06.15 23:13. ╟руюыютюъ: ╩рЁҐшэър эґцфрхҐё т..
╩рЁҐшэър эґцфрхҐё т ъюЁЁхъІшш. ┬Ёюфх с√ ¤ҐюҐ E-V13 чрcхъыш т ╚ёярэшш 7000 ыхҐ эрчрф (ърЁфшры№эр ъхЁрьшър тЁюфх).  | |
|
|
|
| |
╧юёҐ N: 649
╟рЁхушёҐЁшЁютрэ: 24.02.13
╟рьхірэш :
|
|
╬ҐяЁртыхэю: 20.06.15 23:28. ╟руюыютюъ: ┴рҐръ яшЇхҐ: ┬Ёюфх ..
┴рҐръ яшЇхҐ: | ІшҐрҐр: | | ┬Ёюфх с√ ¤ҐюҐ E-V13 чрcхъыш т ╚ёярэшш 7000 ыхҐ эрчрф (ърЁфшры№эр ъхЁрьшър тЁюфх). |
|
╩ґы№ҐґЁр ърЁфшры№эющ ъхЁрьшъш чрЁюфшырё№ эр тюёҐюъх, эр ┴рыърэрє ш юҐҐґфр яЁюэшъыр эр чрярф. ╩ ёюцрыхэш■ яю фЁхтэшь тЁхьхэрь шэЄюЁьрІш юіхэ№ юҐЁ√тюіэр. | |
|
|
|
| |
╧юёҐ N: 848
╟рЁхушёҐЁшЁютрэ: 24.02.13
╟рьхірэш :
|
|
╬ҐяЁртыхэю: 21.03.16 00:00. ╟руюыютюъ: http://cdn.eupedia.c..
 I1b (Z131) ╬фэр шч фтґє тхҐтхщ, ьрыюішёыхээр т ╤хтхЁэ√є └ы№ярє I1a (DF29) ╠эюуюішёыхээр , ЁрёярфрхҐё іхҐ√Ёх тхҐтш, ыюърышчютрээ√х ┬ ╤хтхЁэющ ш ╓хэҐЁры№эющ ┼тЁюях. ╠юцэю ґёыютэю ЁрчфхышҐ№ эр ╠рҐхЁшъют√х ш ╤ърэфшэртёъшх. L22 тхҐт№ яЁхшьґїхёҐтхээю ╤ърэфшэртёър . ─юы I1 т ╪тхІшш ш ═юЁтхушш юъюыю 40% L287 L300 Єшэёъшх ёґсъырф√. ╚ч 28% фюыш I1 т ╘шэы эфшш эр эшє яЁшєюфшҐё 23% ш 5% ёърэфшэртёъшх, Ґюіэхх Їтхфёъшх ёґсъырф√. ╬ёҐры№э√х тхҐтш, ъЁюьх L22, ты ■Ґё ґёыютэю эхьхІъшьш. ╤ююҐтхҐёҐтхээю ¤ъёярэёш эюёшҐхыхщ I1a яЁюшёєюфшыр шч фтґє ¤Ґшє чюэ  | |
|
|
|
| |
═х чрЁхушёҐЁшЁютрэ
╟рЁхушёҐЁшЁютрэ: 01.01.70
|
|
╬ҐяЁртыхэю: 21.03.16 17:31. ╟руюыютюъ: ╨хъґрщ яшЇхҐ: тхҐт№..
╨хъґрщ яшЇхҐ: | ІшҐрҐр: | | тхҐт№ яЁхшьґїхёҐтхээю ╤ърэфшэртёър . ─юы I1 т ╪тхІшш ш ═юЁтхушш юъюыю 40% |
|
─юы 15% ш сюыхх чр шёъы■іхэшхь ■цэюую эхьхІъюую ёхуьхэҐр яЁръҐшіхёъш шфхы№эю ыюцшҐё т уЁрэшІ√ ЁрээхёЁхфэхтхъютющ фхЁцрт√ ╚трЁр ┬шфЄрььх. | |
|
|
|
| |
╧юёҐ N: 856
╟рЁхушёҐЁшЁютрэ: 24.02.13
╟рьхірэш :
|
|
╬ҐяЁртыхэю: 31.03.16 00:32. ╟руюыютюъ: N Africa Composite ..
N Africa Composite Morocco AA 338 0.0% 0.3% 0.6% 0.3% 0.0%
N Africa Mozabite Berbers Algeria AA/Berber 67 3.0% 3.0% 0.0% 3.0% 0.0%
N Africa Northern Egyptians Egypt AA/Semitic 49 6.1% 4.1% 2.0% 4.1% 0.0%
N Africa Berbers from Siwa Egypt AA/Berber 93 28.0% 26.9% 1.1% 23.7% 3.2%
N Africa Baharia Egypt AA/Semitic 41 7.3% 4.9% 2.4% 0.0% 4.9%
N Africa Gurna Oasis Egypt AA/Semitic 34 0.0% 0.0% 0.0% 0.0% 0.0%
N Africa Southern Egyptians Egypt AA/Semitic 69 5.8% 5.8% 0.0% 2.9% 2.9%
C Africa Songhai Niger NS/Songhai 10 0.0% 0.0% 0.0% 0.0% 0.0%
C Africa Fulbe Niger NC/Atlantic 7 14.3% 14.3% 0.0% 14.3% 0.0%
C Africa Tuareg Niger AA/Berber 22 4.5% 4.5% 0.0% 4.5% 0.0%
C Africa Ngambai Chad NS/Sudanic 11 9.1% 9.1% 0.0% 9.1% 0.0%
C Africa Hausa Nigeria (North) AA/Chadic 10 20.0% 20.0% 0.0% 20.0% 0.0%
C Africa Fulbe Nigeria (North) NC/Atlantic 32 0.0% 0.0% 0.0% 0.0% 0.0%
C Africa Yoruba Nigeria (South) NC/Defoid 21 4.8% 4.8% 0.0% 4.8% 0.0%
C Africa Ouldeme Cameroon (Nth) AA/Chadic 22 95.5% 95.5% 0.0% 95.5% 0.0%
C Africa Mada Cameroon (Nth) AA/Chadic 17 82.4% 82.4% 0.0% 76.5% 5.9%
C Africa Mafa Cameroon (Nth) AA/Chadic 8 87.5% 87.5% 0.0% 25.0% 62.5%
C Africa Guiziga Cameroon (Nth) AA/Chadic 9 77.8% 77.8% 0.0% 22.2% 55.6%
C Africa Daba Cameroon (Nth) AA/Chadic 19 42.1% 42.1% 0.0% 36.8% 5.3%
C Africa Guidar Cameroon (Nth) AA/Chadic 9 66.7% 66.7% 0.0% 22.2% 44.4%
C Africa Massa Cameroon (Nth) AA/Chadic 7 28.6% 28.6% 0.0% 14.3% 14.3%
C Africa Other Chadic Cam AA/Chadic 4 75.0% 75.0% 0.0% 25.0% 50.0%
C Africa Shuwa Arabs Cameroon AA/Semitic 5 40.0% 40.0% 0.0% 40.0% 0.0%
C Africa Kanuri Cameroon (Nth) NS/Saharan 7 14.3% 14.3% 0.0% 14.3% 0.0%
C Africa Fulbe Cameroon (Nth) NC/Atlantic 18 11.1% 11.1% 0.0% 5.6% 5.6%
C Africa Moundang Cameroon (Nth) NC/Adamawa 21 66.7% 66.7% 0.0% 14.3% 52.4%
C Africa Fali Cameroon (Nth) NC/Adamawa 48 20.8% 20.8% 0.0% 10.4% 10.4%
C Africa Tali Cameroon (Nth) NC/Adamawa 22 9.1% 9.1% 0.0% 4.5% 4.5%
C Africa Mboum Cameroon (Nth) NC/Adamawa 9 0.0% 0.0% 0.0% 0.0% 0.0%
C Africa Composite Cameroon (Sth) NC/Bantu 90 0.0% 1.1% 0.0% 1.1% 0.0%
C Africa Biaka Pygmies CAR NC/Bantu 33 0.0% 0.0% 0.0% 0.0% 0.0%
W Africa Composite Ч 123 0.0% 0.0% 0.0% 0.0% 0.0%
E Africa Composite Ч 442 0.0% 0.0% 0.0% 0.0% 0.0%
S Africa Composite Ч 105 0.0% 0.0% 0.0% 0.0% 0.0%
| |
|
|
|
| |
╧юёҐ N: 857
╟рЁхушёҐЁшЁютрэ: 24.02.13
╟рьхірэш :
|
|
╬ҐяЁртыхэю: 31.03.16 01:03. ╟руюыютюъ: R1b1c (R-V88) Ouldem..
R1b1c (R-V88)
Ouldeme Cameroon (Nth) AA/Chadic 22 95.5%
Mafa Cameroon (Nth) AA/Chadic 8 87.5%
Mada Cameroon (Nth) AA/Chadic 17 82.4%
Guiziga Cameroon (Nth) AA/Chadic 9 77.8%
Guidar Cameroon (Nth) AA/Chadic 9 66.7%
Moundang Cameroon (Nth) NC/Adamawa 21 66.7%
Daba Cameroon (Nth) AA/Chadic 19 42.1%
Shuwa Arabs Cameroon AA/Semitic 5 40.0%
Massa Cameroon (Nth) AA/Chadic 7 28.6%
Berbers from Siwa Egypt AA/Berber 93 28.0% 26.9%
Fali Cameroon (Nth) NC/Adamawa 48 20.8%
Hausa Nigeria (North) AA/Chadic 10 20.0%
Kanuri Cameroon (Nth) NS/Saharan 7 14.3%
Fulbe Niger NC/Atlantic 7 14.3%
Fulbe Cameroon (Nth) NC/Atlantic 18 11.1%
Tali Cameroon (Nth) NC/Adamawa 22 9.1%
Ngambai Chad NS/Sudanic 11 9.1% 9.1%
Southern Egyptians Egypt AA/Semitic 69 5.8%
Baharia Egypt AA/Semitic 41 7.3% 4.9%
Yoruba Nigeria (South) NC/Defoid 21 4.8%
Tuareg Niger AA/Berber 22 4.5% 4.5%
Northern Egyptians Egypt AA/Semitic 49 6.1% 4.1%
Mozabite Berbers Algeria AA/Berber 67 3.0%
┴юы№ЇшэёҐтю эюёшҐхыхщ R1b т └ЄЁшъх юҐэюёшҐё ъ ьхёҐэюьґ ёґсъырфґ R1b1c V88
═ршсюы№Їр ъюэІхэҐЁрІш т ╤хтхЁэюь ╩рьхЁґэх.
═хсюы№Їюх ъюышіхёҐтю т√єюфІхт шч ┼тЁюя√ ё ёґсъырфюь R1b1a P297 ты ■Ґё яючфэшьш яЁшЇхы№Ірьш, ЁхэхурҐрьш ёЁхфэхтхъют№ .
| |
|
|
|
| |
╧юёҐ N: 858
╟рЁхушёҐЁшЁютрэ: 24.02.13
╟рьхірэш :
|
|
╬ҐяЁртыхэю: 04.04.16 02:00. ╟руюыютюъ: ╚чьхэхэшх сґъхҐр уря..
╚чьхэхэшх сґъхҐр уряыюуЁґяя ёґяхЁ¤Ґэюёр т яЁюІхёёх хую ЁрчтшҐш чртшёшҐ юҐ ¤Ґря𠤥эюухэхчр.
╚эъґсрІшюээ√щ яхЁшюф.
┬ ¤яшІхэҐЁх ¤Ґэюухэхчр шчэріры№эю эрышіхёҐтґхҐ ыюъры№э√щ трЁшрэҐ сґъхҐр уряыюуЁґяя, ъюҐюЁ√щ ьюцхҐ ёґїхёҐтхээю юҐышірҐ№ё юҐ ёЁхфэхёҐрҐшёҐшіхёъюую яю юъЁґцр■їхщ ¤ҐэюышэутшёҐшіхёъющ эхяЁхЁ√тэюёҐш. ╚ыш т юъЁхёҐэюёҐ є ¤яшІхэҐЁр ¤Ґэюухэхчр ьюцхҐ с√Ґ№ ЁрчэюЁюфэр
┬ яЁюІхёёх ёыюцхэш чрЁюф√Їр ёґяхЁ¤Ґэюёр шч ¤Ґюую ыюъры№эюую эрсюЁр яЁюшёєюфшҐ сюыхх ґчър т√сюЁър. ┬ё╕ ¤Ґю юёыюцэ хҐё чэрішҐхы№э√ь тючЁрёҐрэшхь ъюышіхёҐтр ьґҐрІшщ.
┬ юс√іэющ юсёҐрэютъх ьґҐрІш ¤Ґю ёыґірщэюх юҐъыюэхэшх, эх шьх■їхх ъръшє ышсю чэрішҐхы№э√є яюёыхфёҐтшщ.
┬ ёюіхҐрэшш ё т√ёюъшь ґЁютэхь ярёёшюэрЁэюёҐш ¤Ґю ёхЁ№╕чэр чр тър эр ґёяхє.
╧Ёш эшчъющ яыюҐэюёҐш эрёхыхэш т ¤яшІхэҐЁх ¤Ґэюухэхчр яюсхфшҐхыхь ьюцхҐ т√щҐш ш хфшэшіэ√щ яЁхфёҐртшҐхы№ ьґцёъюую яюыр. ╤ т√ёюъющ яыюҐэюёҐ№■ тючэшърхҐ ґцх эхёъюы№ъю ыюъры№э√є уряыюҐшяют.
| |
|
|
|
| |
╧юёҐ N: 859
╟рЁхушёҐЁшЁютрэ: 24.02.13
╟рьхірэш :
|
|
╬ҐяЁртыхэю: 04.04.16 18:22. ╟руюыютюъ: ┬ёяыхёъ ¤эюухэхчр юс..
┬ёяыхёъ ¤эюухэхчр юс√іэю ёюяЁютюцфрхҐё яхЁхєюфюь іхЁхч сґҐ√ыюіэюх уюЁы√Їъю. ╤Ґхяхэ№ ґчюёҐш ьюцхҐ с√Ґ№ Ёрчэр . ╧Ёш эшчъющ яыюҐэюёҐш эрёхыхэш ьюцхҐ яЁюшчющҐш яЁхёхіхэшх тёхє яЁхф√фґїшє ышэшщ ш т√цштрэшх юфэющ хфшэёҐтхээющ ъ Ґюьґ цх яюфтхЁуЇхщё ьґҐрІшш.
└эрышч ¤тюы■Ішюээюую фхЁхтр уряыюуЁґяя ╙ єЁюьюёюь√ яюърч√трхҐ ьэюцхёҐтю Ґръшє ёыґірхт.
┬хҐт№ ┬╥.
╟р яЁхфхы√ └ЄЁшъш т√сЁрырё№ тючьюцэю эхсюы№Їр яюяґы Іш эюёшҐхыхщ ┬╥. ╬ёҐртЇшхё эр "╚ёҐюЁшіхёъющ ╨юфшэх" яЁюЇыш іхЁхч сґҐ√ыюіэюх уюЁы√Їъю ш хфшэёҐтхээ√щ яюҐюьюъ юёҐртшы яюёых ёхс эрсюЁ ьґҐрІшщ тхҐтш ┬. ┬ └чшш Ґръющ цх хфшэёҐтхээ√щ яюҐюьюъ юёҐртшы эрсюЁ ьґҐрІшщ ╤╥.
╟фхё№ юэш Ёрёёхышышё№ тфюы№ яюсхЁхц№ └чшш ш ╬ъхрэшш. ▌Ґю ¤ЄЄхъҐ юёэютрҐхы . ╬фэръю схч ¤ЄЄхъҐр сґҐ√ыюіэюую уюЁы√Їър эх юсюЇыюё№.
┴√ыю Ґръшє яхЁхєюфют сюыхх іхь яЁхфюёҐрҐюіэю. ─ы эрірыр юҐфхышышё№ тхҐтш ╤F ш ED. ╩юҐюЁ√х т ётю■ юіхЁхф№ Ёрёярышё№ эр тхҐтш ╤, F, E, D. ╧Ёш ¤Ґюь тхҐт№ ┼ сыруюяюыґіэю тхЁэґырё№ т └ЄЁшъґ ш шчЁ фэю яюҐхёэшыр ъюЁхээ√х тхҐтш └ ш ┬.
┬ яюёыхфґ■їхь Ґръшх хфшэшіэ√х яхЁхєюф√ ъ эют√ь тхҐт ь с√ыш Ґръ цх эх ЁхфъюёҐ№.
| |
|
|
|
| |
╧юёҐ N: 866
╟рЁхушёҐЁшЁютрэ: 24.02.13
╟рьхірэш :
|
|
╬ҐяЁртыхэю: 10.05.16 13:44. ╟руюыютюъ: └ ш ┬ ╩ръ Ёрч ёЁхфш..
└ ш ┬ ╩ръ Ёрч ёЁхфш сґЇьхэют уряыюуЁґяяр ┬ тёҐЁхірхҐё юҐэюёшҐхы№эю Ёхфъю. ┬ яюёыхфэхщ яю тЁхьхэш ёҐрҐ№х T. Naidoo et al. Development of a single base extension method to resolve Y chromosome haplogroups in sub-Saharan African populations Investig Genet. 2010; 1: 6 хёҐ№ ёҐрҐшёҐшър яю 183 сґЇьхэрь ▐цэющ └ЄЁшъш (Table 3). ─хҐрыш ёсюЁр юсЁрчІют ш яыхьхээр яЁшэрфыхцэюёҐ№ шє эюёшҐхыхщ эх ґҐюіэ ■Ґё , эю, шёєюф шч ёютЁхьхээющ ішёыхээюёҐш ¤Ґюую эрЁюфр (юъюыю 100 Ґ√ё. іхыютхъ), Ґрър т√сюЁър ёююҐтхҐёҐтґхҐ яЁшьхЁэю юфэюьґ юсЁрчІґ эр 275 іхыютхъ. ╥ю хёҐ№, яюъЁ√Ґшх юіхэ№ яыюҐэюх. ╚ч ¤Ґшє 183 іхыютхъ 52 (28,4 %) фрыш яюыюцшҐхы№э√щ ҐхёҐ эр ёэшя ╠51. ▌Ґю A1b1b2a т Ґхъґїхщ тхЁёшш ISOGG. ╩ эшь ьюцэю фюсртшҐ№ эюёшҐхыхщ ёэшяют ╠14 (A1b1a1a) - 5 іхы., M114 (A1b1a1a1a) - 8 іхы., ш P28 (A1b1a1a1b) - 16 іхы. ╚Ґюую, яЁхфёҐртшҐхыш уряыюуЁґяя, юс·хфшэхээ√є яюф A1b1, ёюсшЁр■Ґ 44.3 % сґЇьхэют шч т√сюЁъш. ╙ шє сышцрщЇшє ёюёхфхщ - срэҐґ ▐цэющ └ЄЁшъш, т ¤Ґюь цх шёёыхфютрэшш эрЇыш 5 % (17 шч 343) яЁхфёҐртшҐхыхщ уряыюуЁґяя√ A1b1b2a. ╬іхтшфэю, ёыхфёҐтшх ьхҐшёрІшш ш рёёшьшы Ішш, іҐю, эхёюьэхээю, шьхыр ьхёҐю чр фюыуюх тЁхь ёюёхфёҐтр. ─юёҐрҐюіэю тчуы эґҐ№ эр яюЁҐЁхҐ ═. ╠рэфхы√, ъюёр (xhosa, яхЁт√щ чтґъ - їхыіюъ) яю эрІшюэры№эюёҐш. ═р тҐюЁюь ьхёҐх шфґҐ Ёрчышіэ√х ёґсъырф√, эшёєюф їшх ъ E1b1a: ╠2 (E1b1a) - 24 іхы., M58 (E1b1a1a1a) - 2 іхы., M154 (E1b1a1a1g1c) - 2 іхы., ╠191 (E1b1a1a1f1a) - 14 іхы. ┬ёх тьхёҐх юэш эрсшЁр■Ґ 23,0 % юҐ т√сюЁъш. ╤ сюы№Їющ фюыхщ ґтхЁхээюёҐш яЁюшёєюцфхэшх ¤Ґшє ышэшщ ьюцэю ёт ч𥹠ё ьхҐшёрІшхщ сґЇьхэют юъЁґцртЇшьш шє, ш эрьэюую сюыхх яЁхтюёєюф їшьш яю ішёыхээюёҐш срэҐґ. ╤Ёхфш эшє ¤ҐюҐ цх эрсюЁ ёэшяют фры 66 %. ═р ҐЁхҐ№хь - 29 іхыютхъ (15,8 %) ёю ёэшяюь M35. ▌Ґю уряыюуЁґяяр E1b1b1, єрЁръҐхЁэр фы цшҐхыхщ ╤ЁхфшчхьэюьюЁ№ ш └ЄЁшърэёъюую ╨юур, эю тхё№ьр Ёхфър т ёґсёрєрЁёъющ └ЄЁшъх (1,5 % ґ срэҐґ ▐цэющ └ЄЁшъш т Ґюь цх шёёыхфютрэшш). ╦шЇ№ чрҐхь ёыхфґ■Ґ 13 іхыютхъ (7,1 %) ёю ёэшяюь ╨6 (B2b1), ъ ъюҐюЁ√ь яЁшь√ър■Ґ юсырфрҐхыш ёэшяют M112 (B2b*) - 2 іхы., ш ╨8 (B2b4a) - 5 іхы. ╚Ґюую яюыґірхҐё 10,9 % яЁхфёҐртшҐхыхщ уряыюуЁґяя√ B2b, яЁръҐшіхёъш юҐёґҐёҐтґ■їхщ т т√сюЁъх ■цэюрЄЁшърэёъшє срэҐґ (1 шч 343). ╤Ёхфш яюёыхфэшє ё ірёҐюҐющ 16 % тёҐЁхірхҐё фЁґующ ёґсъырф - B2a1a (M152). ╚ч юёҐртЇшхё 10-Ґш іхыютхъ 6 шыш 7 - тэ√х яюҐюьъш хтЁюяхщёъшє яюёхыхэІхт шч уряыюуЁґяя I (M170) - 1, R1a1a (M198) - 2, ш R1b (M343) - 3. 300 ыхҐ ёютьхёҐэюую яЁюцштрэш ш ЁрёяЁюёҐЁрэхээр яЁръҐшър сґЁёъшє ЄхЁьхЁют сЁрҐ№ ёхсх эрыюцэшІ шч ьхёҐэюую эрёхыхэш эх ьюуыш эх ёърч𥹸 . ╬фшэ эюёшҐхы№ ёэшяр M34 (E1b1b1b2a1) ьюцхҐ, т яЁшэІшях, юърч𥹸 ъръ яюҐюьъюь схыюую шььшуЁрэҐр, Ґръ ш юфэшь шч іыхэют тхҐтш, ьрЁъшЁютрээющ ╠35. ═ръюэхІ, ҐЁюх - яЁхфёҐртшҐхыш уряыюуЁґяя√ ┼2, тёҐЁхір■їхщё ё ірёҐюҐющ 10,2 % ґ ■цэюрЄЁшърэёъшє срэҐґ. ╬іхтшфэю, юҐ яюёыхфэшє ш фюёҐрырё№. ╚Ґюую, ёяхІшЄшіхіхёъш ъющёрэёъшьш ышэш ьш т ¤Ґющ т√сюЁъх ьюцэю эрчт𥹠A1b1, B2b ш E1b1b1, ъюҐюЁ√х тьхёҐх ёюсшЁр■Ґ 71 % т√сюЁъш ■цэюрЄЁшърэёъшє сґЇьхэют. ╧юёыхфэ уЁґяяр фрыр юҐЁшІрҐхы№э√х Ёхчґы№ҐрҐ√ яю эшёєюф їшь ёэшярь M78 (E1b1b1a1), M148 (E1b1b1a1c1), M81 (E1b1b1b1a), M107 (E1b1b1b1a1 - private), M165 (E1b1b1b1a2a - private), M123 (E1b1b1b2a), M34 (E1b1b1b2a1), M136 (E1b1b1b2a1a1) ш M281 (E1b1b2), іҐю ґтюфшҐ хх юсїхую яЁхфър ё хтЁюяхщёъшьш ш сышцэхтюёҐюіэ√ьш тхҐт ьш Ґюую цх ёґсъырфр, ъръ ьшэшьґь, эр 12000 ыхҐ эрчрф. ┬ ¤Ґющ тхё№ьр ЁхяЁхчхэҐрҐштэющ т√сюЁъх (ёь. т√Їх) эх с√ыш юсэрЁґцхэ√ уряыюҐшя√ └0, іҐю ёҐртшҐ яюф ёюьэхэшх яюяґы Ёэюх ґҐтхЁцфхэшх, іҐю сґЇьхэ√ - яюҐюьъш ёрь√є Ёрээшє ы■фхщ. ╚є эхюс√ірщэю сюы№Їр трЁшрҐштэюёҐ№ - ёыхфёҐтшх эрыюцхэш эхёъюы№ъшє фрыхъю ЁрчюЇхфЇшєё ышэшщ, яЁюшчтюфэ√є юҐ ры№Єр-уряыюуЁґяя√, р тютёх эх чэръ Ґюую, іҐю ¤Ґр ¤Ґэшіхёър уЁґяяр ърър -Ґю юёюсхээю фЁхтэ . ├фх ш ъюуфр юэш яхЁхьхЇрышё№, фрт т шҐюух ёютЁхьхээґ■ яюяґы Іш■ ъющёрэёъшє эрЁюфют, ь√ эх чэрхь. ┼ёыш шёєюфшҐ№ шч Ґюую, іҐю уряыюЁґяя√ A1b1 ш E1b1b1 тхё№ьр єрЁръҐхЁэ√ фы ■ур └Ёртшщёъюую я-тр ш └ЄЁшърэёъюую ╨юур, Ґю яю шьх■їшьё фрээ√ь ьюцэю яЁхфяюыюцшҐ№, іҐю яЁхфъш сґЇьхэют ёЄюЁьшЁютрышё№ уфх-Ґю т ¤Ґюь Ёрщюэх ьхцфґ 15 ш 12 Ґ√ё. ыхҐ эрчрф, ш ышЇ№ чрҐхь ьшуЁшЁютрыш эр ■у └ЄЁшъш. ┬ёҐЁхҐшыш ыш юэш Ґрь фЁґушє ы■фхщ ш ёьхЇрышё№ ё эшьш, шыш Ґх ґцх т√ьхЁыш ъ Ґюьґ тЁхьхэш, ь√ эх чэрхь. ┴хёёяюЁэю ышЇ№, іҐю ч𠤥ющ ьшуЁрІшхщ яюёыхфютры фюыушщ яхЁшюф шчюы Ішш (тяыюҐ№ фю яю тыхэш ёъюҐютюфют срэҐґ т эрірых эрЇхщ ¤Ё√), т Ґхіхэшх ъюҐюЁюую "т√ь√ышё№" фЁґушх ухэхрыюушіхёъшх ышэшш. ┬ ётю■ юіхЁхф№, ёюєЁрэшышё№ Ґх, іҐю юърчрышё№ ьшэюЁэ√ьш тю тЁхьхэр эхюышҐшіхёъющ Ёхтюы■Ішш эр ┴ышцэхь ┬юёҐюъх, ш шёіхчыш Ґрь. ╥ръ іҐю шьх■їшхё фрээ√х яю Y єЁюьюёюьх ъющёрэёъшє эрЁюфют эх фр■Ґ эшъръщ фюяюыэшҐхы№эющ яюффхЁцъш ушяюҐхчх Out-of-Africa. ┬ёх ыюушіэю юс· ёэ хҐё схч яЁштыхіхэш хх яюёҐґырҐют.
| |
|
|
|
| |
╧юёҐ N: 883
╟рЁхушёҐЁшЁютрэ: 24.02.13
╟рьхірэш :
|
|
╬ҐяЁртыхэю: 31.08.16 18:02. ╟руюыютюъ: ┬ эрёҐю їхх тЁхь эх..
┬ эрёҐю їхх тЁхь эхсюы№Їющ яЁюІхэҐ (юҐ 1 фю 4%) шч ╨1с-V88 тёҐЁхір■Ґё т ╦хтрэҐх, ёЁхфш ╦штрэІхт, ─Ёґчют ш хтЁххт, ш яюіҐш т ърцфющ ёҐЁрэх └ЄЁшъш ъ ёхтхЁґ юҐ ¤ътрҐюЁр. ┴юыхх т√ёюър ірёҐюҐр т ┼ушяҐх (5%), ёЁхфш ┴хЁсхЁют юҐ хушяхҐёъю-ыштшщёъющ уЁрэшІ√ (23%), ёЁхфш ╤ґфрэёъшє ъюяҐют (15%), ╒рґёр эрЁюф ╤ґфрэр (эр 40%), ╘ґы№сх цшҐхы ь ╤рєхы (54% т ═шухЁх ш ╩рьхЁґэх), ш Chadic яыхьхэ ╤хтхЁэющ ═шухЁшш ш ╤хтхЁэюь ╩рьхЁґэх (юёюсхээю ёЁхфш ╩шЁфш), уфх юэ эрсы■фрхҐё эр ірёҐюҐх т фшрярчюэх юҐ 30% фю 95% ьґцішэ. ╧ю фрээ√ь Cruciani ёюртҐ. (2010) ╨1с-V88 с√ яхЁхёхъ ╤рєрЁґ ьхцфґ 9,200 ш 5600 ыхҐ эрчрф, ш, тхЁю Ґэю, ёт чрээ√х ё ЁрёяЁюёҐЁрэхэшхь ч√ъют Chadic, Єшышры └ЄЁрчшщёъшх ч√ъш. V88 с√ ьшуЁшЁютрыш шч ┼ушяҐр т ╤ґфрэ, чрҐхь ЁрёЇшЁ хҐё тфюы№ ╤рєхы фю ╤хтхЁэюую ╩рьхЁґэр ш ═шухЁшш. ╬фэръю, ╨1с-V88 эх Ґюы№ъю ёЁхфш Chadic т√ёҐґяртЇшє, эю ш ёЁхфш Senegambian ъюыюэъш (╘ґырэ-╒рґёр) ш ╤хьшҐёъюую ч√ър (┴хЁсхЁ√, рЁрс√).
╨1с-V88 тёҐЁхірхҐё ёЁхфш ъюЁхээюую эрёхыхэш ╨ґрэф√, ▐└╨, ═рьшсш , └эуюыр, ╩юэую, ├рсюэ, ▌ътрҐюЁшры№эр ├тшэх , ╩юҐ-ф'╚тґрЁ, ├тшэх -┴шёрґ. ╪шЁюъюх ЁрёяЁюёҐЁрэхэшх V88 тю тёхє ірёҐ є └ЄЁшъш, хх ЁрёяЁюёҐЁрэхээюёҐш ёЁхфш я𸥴Ї№шє яыхьхэ, ш ъюрыхёІхэІшш тючЁрёҐ уряыюуЁґяя√ тёх яюффхЁцштр■Ґ Ёрчуюэ эхюышҐр. ┬ ы■сюь ёыґірх, сюыхх яючфэ ьшуЁрІш шч ┼ушяҐр с√ыю с√ эхтхЁю Ґэю.
| |
|
|
|
|
|
| |
╧юёҐ N: 979
╟рЁхушёҐЁшЁютрэ: 24.02.13
╟рьхірэш :
|
|
╬ҐяЁртыхэю: 27.07.17 11:25. ╟руюыютюъ: ├ряыюуЁґяя√ эхюышҐші..
├ряыюуЁґяя√ эхюышҐшіхёъюую эрёхыхэш ╪юҐырэфшш (Ґю хёҐ№ яЁхфЇхёҐтхээшъют ъґы№ҐґЁ√ ъюыюъютшфэ√є ъґсъют):
I3049 England_Neolithic Great Britain I
I0518 England_Neolithic Great Britain I2a2
I0519 England_Neolithic Great Britain I2a1b
I0520 England_Neolithic Great Britain I2a2a1
I2631 Scotland_Neolithic Great Britain I2a1b
I2630 Scotland_Neolithic Great Britain I2a1b1
I2932 Scotland_Neolithic Great Britain I2a1b
I2933 Scotland_Neolithic Great Britain I2a2a1a1a2
I2935 Scotland_Neolithic Great Britain I2a1b1
I2635 Scotland_Neolithic Great Britain I2a1b1
I3133 Scotland_Neolithic Great Britain I2a1b1
I3134 Scotland_Neolithic Great Britain I2a2a1a1a
I3135 Scotland_Neolithic Great Britain I2a2a
I3137 Scotland_Neolithic Great Britain I
I2650 Scotland_Neolithic Great Britain I2a2a1b
I2977 Scotland_Neolithic Great Britain I2a2
I2979 Scotland_Neolithic Great Britain I2a1b1
I2629 Scotland_Neolithic Great Britain I2
I2978 Scotland_Neolithic Great Britain I2a1b
I2637 Scotland_Neolithic Great Britain I2a1b
I2660 Scotland_Neolithic Great Britain I2a2a1a1a
I2634 Scotland_Neolithic Great Britain I2a1b
I2796 Scotland_Neolithic Great Britain I2a1b
I2691 Scotland_Neolithic Great Britain I2a2a1
I2657 Scotland_Neolithic Great Britain I2a2a
Bell Beakers:
I1388 BB_Southern_France France R1b1a1a2a1a
I1390 BB_Central_Europe France R1b1a1a2a1a2
I1389 BB_Central_Europe France R1b1a1a2a1a2
I1381 BB_Central_Europe France R1b1a1a2a1a
I1382 BB_Central_Europe France R1b1a1a2a1a2
I3875 BB_Southern_France France R1b1a1a2a1a2b1
I2478 BB_Northern_Italy Italy R1b1a1a2a1a2
I1767 BB_Britain Great Britain I2a2a1a1a
I2453 BB_Britain Great Britain R1b1a1a2a1a2c1
I2452 BB_Britain Great Britain R1b1a1a2a1a2c1
I2445 BB_Britain Great Britain R1b1a1a2a1a2c1
I2447 BB_Britain Great Britain R1b1a1a2a1a2c1
I2416 BB_Britain Great Britain R1b1a1a2a1a
I2565 BB_Britain Great Britain R1b1a1a2a1a2c
I3256 BB_Britain Great Britain R1b1a1a2a1a2c1
I4068 BB_Central_Europe The Netherlands R1b1a1a2a1a2
I4069 BB_Central_Europe The Netherlands R1b1a1a2a1a2
I4070 BB_Central_Europe The Netherlands R1b1a1a2a1a1
I4073 BB_Central_Europe The Netherlands R1b1a1a2a1a2
I4074 BB_Central_Europe The Netherlands R1b1a1a2a1a2
I2364 BB_Central_Europe Hungary H2
I2365 BB_Central_Europe Hungary R1b1a1a2a1a2b1
I2741 BB_Central_Europe Hungary I2a1a1
I2786 BB_Central_Europe Hungary I2a2a
I2787 BB_Central_Europe Hungary R1b1a1a2a2
I4178 BB_Central_Europe Hungary R1b1a1a2
I3528 BB_Central_Europe Hungary G2a2a1a2a1
I4251 BB_Central_Europe Poland R1b1a1a2
I4252 BB_Central_Europe Poland R
I4253 BB_Central_Europe Poland R1b1a1a2
I3604 BB_Central_Europe Germany R1b1a
I3607 BB_Central_Europe Germany R1b1a1a
I3599 BB_Central_Europe Germany R1b1a1a2a1a2
I3588 BB_Central_Europe Germany R1b1a1a2a1a
I3589 BB_Central_Europe Germany R1b1a1a2a1a2b1
| |
|
|
|
| |
╧юёҐ N: 980
╟рЁхушёҐЁшЁютрэ: 24.02.13
╟рьхірэш :
|
|
╬ҐяЁртыхэю: 27.07.17 11:26. ╟руюыютюъ: The Genomic History..
The Genomic History Of Southeastern Europe
I2407 Globular_Amphora_Poland_brother.of.I2435_son.of.I2433_son.of.I2440 Kierzkowo Poland BT
I3498 Starcevo_EN Beli Manastir-Popova zemlja Croatia C
I1296 Balkans_MP_Neolithic Malak Preslavets Bulgaria C
I0706 Balkans_Dzhulyunitsa_Neolithic Dzhulyunitsa Bulgaria C
I5070 LBK_Austria Schletz Austria C1a2
Tep006 Anatolia_Neolithic_Tepecik_Ciftlik.SG Tepecik Ciftlik Turkey C1a2
I0585 Iberia_HG La Brana-Arintero, Leon Spain C1a2
I1496 LBK_Hungary_MN Apc-Berekalya Hungary C1a2
I1500 ALPc_MN Kompolt-Kigyoser Hungary C1a2
I3947 Croatia_Cardial_Neolithic Zemunica Cave Croatia C1a2
Vestonice16 Vestonice16 Dolni Vestonice Czech C1a2
I1102 Anatolia_Neolithic Barcın, Marmara Region, Northwest Anatolia Turkey C1a2
GoyetQ116-1_published GoyetQ116-1_published Goyet Belgium C1a2
I4089 Romania_Chalcolithic Urziceni Romania C1a2a
Kostenki14 Kostenki14 Kostenki Russia C1b
I0795 LBK_EN Karsdorf Germany CT
I0804 Unetice_EBA Eulau Germany CT
I1700 PPNB 'Ain Ghazal Jordan CT
I1727 PPNB 'Ain Ghazal Jordan CT(xE,G,J,LT,R,Q1a,Q1b)
I1416 PPNB 'Ain Ghazal Jordan CT
ANI152 Bulgaria_Varna_Eneolithic1 Varna Bulgaria CT
I2435 Globular_Amphora_Poland_brother.of.I2407_son.of.I2433_son.of.I2440 Kierzkowo Poland CT
I2426 Bulgaria_Late_Chalcolithic2 Sushina Bulgaria CT
I3151 Trypillia Verteba Cave Ukraine E
mota.SG Mota.SG Mota Cave, Gamo Highlands Ethiopia E1b1a2
I1415 PPNB 'Ain Ghazal Jordan E1b1b1
I1710 PPNB 'Ain Ghazal Jordan E1b1b1a
I3948 Croatia_Cardial_Neolithic Zemunica Cave Croatia E1b1b1a1b1
I1414 PPNB 'Ain Ghazal Jordan E1b1b1b2
ANI160 Bulgaria_Varna_Eneolithic2 Varna Bulgaria G2
I0727 Anatolia_Neolithic Mentese Turkey G2a
I2111 Trypillia Verteba Cave Ukraine G2a
I0659 LBK_EN Halberstadt-Sonntagsfeld Germany G2a2a
Tep003 Anatolia_Neolithic_Tepecik_Ciftlik.SG Tepecik Ciftlik Turkey G2a2a
I0048 LBK_EN Halberstadt-Sonntagsfeld Germany G2a2a
I0551 Salzmuende_MN Salzmuende-Schiebzig Germany G2a2a
I0633 Balkans_Neolithic Gomolava, Hrtkovci, Vojvodina Serbia G2a2a1
I0821 LBK_EN Halberstadt-Sonntagsfeld Germany G2a2a1
I0723 Anatolia_Neolithic Mentese Turkey G2a2a1
I0634 Balkans_Neolithic Gomolava, Hrtkovci, Vojvodina Serbia G2a2a1a
I1131 Balkans_Neolithic Gomolava, Hrtkovci, Vojvodina Serbia G2a2a1a
I2510 Bulgaria_BA Dzhulyunitsa Bulgaria G2a2a1a2
Klei10.SG Greece_Final_Neolithic.SG Kletios Greece G2a2a1a2
RISE374.SG Maros.SG Szöreg, Sziv Utca Hungary G2a2a1a2a
I2792 Croatia_Vucedol Vucedol tell Croatia G2a2a1a2a
I1099 Anatolia_Neolithic Barcın, Marmara Region, Northwest Anatolia Turkey G2a2a1a2a
I1103 Anatolia_Neolithic Barcın, Marmara Region, Northwest Anatolia Turkey G2a2a1a2a
I1583 Anatolia_Neolithic Barcın, Marmara Region, Northwest Anatolia Turkey G2a2a1a2a
I0698 Bulgaria_Neolithic Yabalkovo Bulgaria G2a2a1a2a
Iceman.SG Iceman_MN.SG Ötz valley, Tyrol Italy G2a2a1a2a1a
I1295 Balkans_MP_Neolithic Malak Preslavets Bulgaria G2a2b2a
I3879 Balkans_MP_Neolithic Malak Preslavets Bulgaria G2a2b2a
I1926 Trypillia Verteba Cave Ukraine G2a2b2a
I0744 Anatolia_Neolithic Barcın, Marmara Region, Northwest Anatolia Turkey G2a2b2a
I2110 Trypillia Verteba Cave Ukraine G2a2b2a
I1927 Trypillia_outlier Verteba Cave Ukraine G2a2b2a1a1b1a1a1
I0746 Anatolia_Neolithic Barcın, Marmara Region, Northwest Anatolia Turkey G2a2b2a1a1c
I2431 Bulgaria_Middle_Chalcolithic Ivanovo Bulgaria G2a2b2a1a1c1a
S5204.E1.L1 LBK_Austria Schletz Austria G2a2b2a3
I1097 Anatolia_Neolithic Barcın, Marmara Region, Northwest Anatolia Turkey G2a2b2a3
ANI159-ANI181 Bulgaria_Varna_Eneolithic2 Varna Bulgaria G2a2b2b
I2532 Romania_EN Cotatcu Romania G2a2b2b
Bon001 Anatolia_Neolithic_Boncuklu.SG Boncuklu Turkey G2a2b2b
Bon004 Anatolia_Neolithic_Boncuklu.SG Boncuklu Turkey G2a2b2b
I0676 Macedonia_Neolithic Govrlevo, Sopi_te, Skopje Macedonia G2a2b2b1
I2521 Bulgaria_Neolithic Dzhulyunitsa Bulgaria G2a2b2b1a
I1314 Iberia_Chalcolithic El Mirador Cave, Atapuerca, Burgos Spain G2a2b2b1a1
I0745 Anatolia_Neolithic Barcın, Marmara Region, Northwest Anatolia Turkey H2
I0709 Anatolia_Neolithic Barcın, Marmara Region, Northwest Anatolia Turkey H2
I1101 Anatolia_Neolithic Barcın, Marmara Region, Northwest Anatolia Turkey H2
I0405 Iberia_MN La Mina Spain H2
I2520 Bulgaria_BA Dzhulyunitsa Bulgaria H2
I0174 Starcevo_EN Alsonyek-Bataszek Hungary H2
I0867 PPNB Motza Israel H2
RISE489.SG Remedello_BA.SG Remedello di Sotto Italy I
RISE175.SG Nordic_BA.SG Abekås Sweden I
I0724 Anatolia_Neolithic Mentese Turkey I
S5886.E1.L1 Ukraine_Neolithic Dereivka Ukraine I
Chaudardes1_m Chaudardes1 Chaudardes France I
BerryAuBac BerryAuBac Berry au bac France I
RISE486.SG Remedello_BA.SG Remedello di Sotto Italy I
I0807 Baalberge_MN Esperstedt Germany I
RISE487.SG Remedello_BA.SG Remedello di Sotto Italy I
Rochedane Rochedane Rochedane France I
I1282 Iberia_Chalcolithic El Mirador Cave, Atapuerca, Burgos Spain I
RISE210.SG Nordic_BA.SG Ängamöllan Sweden I
I1284 Iberia_Chalcolithic El Mirador Cave, Atapuerca, Burgos Spain I
S5771.E1.L1 Iron_Gates_HG Vlasac Serbia I
RISE179.SG Nordic_LN.SG Abekås Sweden I
S5773.E1.L1 Iron_Gates_HG Vlasac Serbia I
RISE254.SG Vatya.SG Százhalombatta-Földvár Hungary I
I3713 Ukraine_Neolithic Volniensky, Vilnianka Ukraine I
S5869.E1.L1 Ukraine_Neolithic_1d_relative_of_S5870.E1.L1 Volniensky, Vilnianka Ukraine I
S5872.E1.L1 Ukraine_Neolithic Volniensky, Vilnianka Ukraine I
S5868.E1.L1 Ukraine_Neolithic Volniensky, Vilnianka Ukraine I
I4167 Sopot_LN Beli Manastir-Popova zemlja Croatia I
Bul8 Bulgaria_Beli_Breyag_EBA Beli Breyag Bulgaria I
I2440 Globular_Amphora_Poland Kierzkowo Poland I2
I3716 Ukraine_Neolithic Volniensky, Vilnianka Ukraine I2
I4607 Iron_Gates_HG Schela Cladovei Romania I2
I4870 Iron_Gates_HG Vlasac Serbia I2
I4440 Latvia_HG Zvejnieki Latvia I2a1
I4438 Latvia_HG Zvejnieki Latvia I2a1
I5236 Iron_Gates_HG Padina Serbia I2a1
I1495 Lengyel_LN Apc-Berekalya Hungary I2a1
I0015 Motala_HG Motala, Kanaljorden Sweden I2a1
I1507 Koros_Hungary_EN_HG Tiszaszolos-Domahaza Hungary I2a1
I1763 Ukraine_Mesolithic Vasil'evka Ukraine I2a1
I1303 Iberia_Chalcolithic El Mirador Cave, Atapuerca, Burgos Spain I2a1a1a
Bichon.SG Bichon Grotte du Bichon Switzerland I2a1a2a
Loschbour_published.DG Loschbour_published.DG Echternach Luxembourg I2a1b
I0013 Motala_HG Motala, Kanaljorden Sweden I2a1b
I0412 Iberia_EN Els Trocs Spain I2a1b1
I0172 Esperstedt_MN Esperstedt Germany I2a1b1a1
I0017 Motala_HG Motala, Kanaljorden Sweden I2a1b2a1
I2403 Globular_Amphora_Poland Kierzkowo Poland I2a2
S5870.E1.L1 Ukraine_Neolithic Volniensky, Vilnianka Ukraine I2a2
I2529 Bulgaria_Neolithic Yabalkovo Bulgaria I2a2
I4915 Iron_Gates_HG Hadučka Vodenica Serbia I2a2
S5888.E1.L1 Ukraine_Neolithic_father_or_son_of_S5875.E1.L1 Dereivka Ukraine I2a2
Bul6 Bulgaria_Beli_Breyag_EBA Beli Breyag Bulgaria I2a2
I2405 Globular_Amphora_Poland Kierzkowo Poland I2a2
I3714 Ukraine_Neolithic Volniensky, Vilnianka Ukraine I2a2a
I4878 Iron_Gates_HG Vlasac Serbia I2a2a
Falkenstein Falkenstein Falkenstein-Höhle, Swabian Jura, Baden-Württemberg, Germany I2a2a
RISE247.SG Vatya.SG Százhalombatta-Földvár Hungary I2a2a1
I4551 Latvia_HG Zvejnieki Latvia I2a2a1
I4553 Latvia_HG Zvejnieki Latvia I2a2a1
RISE479.SG Vatya.SG Erd Hungary I2a2a1a2a
I2441 Globular_Amphora_Poland Kierzkowo Poland I2a2a1b
I4596 Latvia_HG Zvejnieki Latvia I2a2a1b
I4441 Latvia_HG Zvejnieki Latvia I2a2a1b
ILK001 Globular_Amphora_Ukraine Ilyatka Ukraine I2a2a1b
I2176 Bulgaria_EBA Smyadovo Bulgaria I2a2a1b
I4882 Iron_Gates_HG_brother_of_I4880 Vlasac Serbia I2a2a1b
S5875.E1.L1 Ukraine_Neolithic Dereivka Ukraine I2a2a1b
I3715 Ukraine_Neolithic Volniensky, Vilnianka Ukraine I2a2a1b1
I3717 Ukraine_Neolithic Dereivka Ukraine I2a2a1b1
I2175 Bulgaria_EBA Smyadovo Bulgaria I2a2a1b1
Bul4 Yamnaya_Bulgaria_outlier Mednikarovo Bulgaria I2a2a1b1b
I1738 Ukraine_Neolithic Vovnigi Ukraine I2a2a1b1b
I2165 Bulgaria_EBA Merichleri, Kairyaka necropolis Bulgaria I2a2a1b1b
RISE552.SG Yamnaya_Kalmykia.SG Ulan Russia I2a2a1b1b2
I0406 Iberia_MN La Mina Spain I2a2a1b2
I4880 Iron_Gates_HG Vlasac Serbia I2a2a1b2
ILK002 Globular_Amphora_Ukraine Ilyatka Ukraine I2a2a1b2
I4881 Iron_Gates_HG Vlasac Serbia I2a2a1b2
I4914 Iron_Gates_HG Hadučka Vodenica Serbia I2a2a1b2
I1277 Iberia_Chalcolithic El Mirador Cave, Atapuerca, Burgos Spain I2a2a2a
I0012 Motala_HG Motala, Kanaljorden Sweden I2c
I1096 Anatolia_Neolithic Barcın, Marmara Region, Northwest Anatolia Turkey I2c
I0116 Unetice_EBA Esperstedt Germany I2c
I3712 Ukraine_Neolithic Volniensky, Vilnianka Ukraine IJ
I0211 Karelia_HG Yuzhnyy Oleni Ostrov, Karelia Russia J
Iboussieres25-1 Iboussieres25-1 Aven des Iboussières à Malataverne, Rhône-Alpes France J?
SATP.SG Satsurblia Satsurblia Georgia J1
I2495 Anatolia_BA Harmanoren-Gondurle Hoyuk, Isparta Province, Western Turkey Turkey J1a
I5068 LBK_Austria Kleinhadersdorf Flur Marchleiten Austria J2
KK1.SG Kotias Kotias Klde Georgia J2a
I0708 Anatolia_Neolithic Barcın, Marmara Region, Northwest Anatolia Turkey J2a
I1504 Hungary_LBA Ludas-Varju-Dulo Hungary J2a1
I4331 Croatia_EMBA Veliki Vanik Croatia J2b2a
I1709 PPNB 'Ain Ghazal Jordan nodata
I0434 Samara_Eneolithic Khvalynsk, Volga River, Samara Russia Q1a
I4550 Latvia_HG Zvejnieki Latvia Q1a2
MA1.SG MA1_HG.SG Mal'ta Russia R
I1949 Ganj_Dareh_Iran_Neolithic Ganj Dareh Iran R
I1542 Corded_Ware_Germany Esperstedt Germany R
Iboussieres31-2 Iboussieres31-2 Aven des Iboussières à Malataverne, Rhône-Alpes France R
I1534 Corded_Ware_Germany Esperstedt Germany R
S5891.E1.L1 Ukraine_Neolithic Dereivka Ukraine R
I1945 Ganj_Dareh_Iran_Neolithic Ganj Dareh Iran R
I4112 Ukraine_Neolithic Dereivka Ukraine R
I4655 Iron_Gates_HG Schela Cladovei Romania R
I2181 Bulgaria_Late_Chalcolithic1 Smyadovo Bulgaria R
I0246 Potapovka Utyevka, Samara River, Samara Russia R1
S5878.E1.L1 Ukraine_Neolithic_father_or_son_of_S5883.E1.L1 Dereivka Ukraine R1
RISE434.SG Corded_Ware_Germany.SG Tiefbrunn Germany R1
S5881.E1.L1 Ukraine_Neolithic Dereivka Ukraine R1
ANI153 Bulgaria_Varna_Eneolithic1 Varna Bulgaria R1
I1536 Corded_Ware_Germany Esperstedt Germany R1a
I1538 Corded_Ware_Germany Esperstedt Germany R1a
RISE386.SG Sintashta_MBA_RISE.SG Bulanovo Russia R1a
I1540 Corded_Ware_Germany Esperstedt Germany R1a1
S5876.E1.L1 Ukraine_Neolithic Dereivka Ukraine R1a
I1819 Ukraine_Mesolithic Vasil'evka Ukraine R1a
I1544 Corded_Ware_Germany Esperstedt Germany R1a
I0433 Samara_Eneolithic Khvalynsk, Volga River, Samara Russia R1a1
I0061 Karelia_HG Yuzhnyy Oleni Ostrov, Karelia Russia R1a1a1
I0361 Srubnaya Spiridonovka IV, Samara River, Samara Russia R1a1a
I1532 Corded_Ware_Germany Esperstedt Germany R1a1a
RISE446.SG Corded_Ware_Germany.SG Bergrheinfeld Germany R1a1a1
I0104 Corded_Ware_Germany Esperstedt Germany R1a1a1
RISE61.SG Nordic_MN_B.SG Kyndelöse Denmark R1a1a1
RISE94.SG BattleAxe_Sweden.SG Viby Sweden R1a1a1
RISE512.SG Andronovo.SG Kytmanovo Russia R1a1a1
RISE392.SG Sintashta_MBA_RISE.SG Stepnoe Cemetery Russia R1a1a1b
I0419 Potapovka Utyevka, Samara River, Samara Russia R1a1a1b
I0099 Halberstadt_LBA Halberstadt-Sonntagsfeld Germany R1a1a1b1a2
I0424 Srubnaya Uvarovka, Samara River, Samara Russia R1a1a1b2
I0232 Srubnaya Novoselki, Northern Forest, Samara Russia R1a1a1b2
I2163 Bulgaria_MLBA Merichleri, Kairyaka necropolis Bulgaria R1a1a1b2
I0423 Srubnaya_Outlier Barinovka, Samara River, Samara Russia R1a1a1b2
I0432 Poltavka_outlier Sok River, Samara Russia R1a1a1b2a
I0430 Srubnaya Spiridonovka II, Samara River, Samara Russia R1a1a1b2a2a
I0247 Scythian_IA Nadezhdinka, Volga Steppes, Samara Russia R1a1a1b2a2a
S5879.E1.L1 Ukraine_Neolithic_brother_of_S5890.E1.L1_father_or_son_of_I3718 Dereivka Ukraine R1b1(xR1b1a1a2)
RISE436.SG Corded_Ware_Germany.SG Tiefbrunn Germany R1b1
S5893.E1.L1 Ukraine_Neolithic Dereivka Ukraine R1b1
I0410 Iberia_EN Els Trocs Spain R1b1a(xR1b1a1a2)
I5235 Iron_Gates_HG Padina Serbia R1b1a(xR1b1a1a,xR1b1a1a2)
I4114 Ukraine_Neolithic Dereivka Ukraine R1b1a
I5232 Iron_Gates_HG_outlier Padina Serbia R1b1a(xR1b1a1a,xR1b1a1a2)
I5237 Iron_Gates_HG Padina Serbia R1b1a(xR1b1a1a,xR1b1a1a2)
Villabruna Villabruna Villabruna Italy R1b1a
I5240 Iron_Gates_HG Padina Serbia R1b1a(xR1b1a1a,xR1b1a1a2)
I4081 Iron_Gates_HG Ostrovul Corbului Romania R1b1a
I0122 Samara_Eneolithic Khvalynsk, Volga River, Samara Russia R1b1a
I4916 Iron_Gates_HG Hadučka Vodenica Serbia R1b1a(xR1b1a1,xR1b1a1a,xR1b1a1a2)
I2430 Bulgaria_Late_Chalcolithic Smyadovo Bulgaria R1b1a
I3718 Ukraine_Neolithic Dereivka Ukraine R1b1a(xR1b1a1a,xR1b1a1a2)
S5890.E1.L1 Ukraine_Neolithic Dereivka Ukraine R1b1a(xR1b1a1a,xR1b1a1a2)
I4666 Lepenski_Vir_Neolithic Lepenski Vir Serbia R1b1a
S5883.E1.L1 Ukraine_Neolithic Dereivka Ukraine R1b1a(xR1b1a1a,xR1b1a1a2)
I0559 Baalberge_MN Quedlinburg Germany R1b1a(xR1b1a1a2)
S5772.E1.L1 Iron_Gates_HG Vlasac Serbia R1b1a(xR1b1a1a,xR1b1a1a2)
RISE431.SG Corded_Ware_Proto_Unetice_Poland.SG Leki Male Poland R1b1a
S5892.E1.L1 Ukraine_Neolithic Dereivka Ukraine R1b1a
I0124 Samara_HG Lebyanzhinka, Sok River, Samara Russia R1b1a1a
I4630 Latvia_HG Zvejnieki Latvia R1b1a1a(xR1b1a1a2)
I4626 Latvia_EN Zvejnieki Latvia R1b1a1a
I4439 Latvia_HG Zvejnieki Latvia R1b1a1a(xR1b1a1a2)
I4432 Latvia_HG Zvejnieki Latvia R1b1a1a(xR1b1a1a2)
I4628 Latvia_EN Zvejnieki Latvia R1b1a1a(xR1b1a1a2)
I4436 Latvia_HG Zvejnieki Latvia R1b1a1a(xR1b1a1a2)
I4627 Latvia_MN Zvejnieki Latvia R1b1a1a(xR1b1a1a2)
I4434 Latvia_HG Zvejnieki Latvia R1b1a1a(xR1b1a1a2)
I0371 Poltavka Grachevka, Sok River, Samara Russia R1b1a1a2
RISE550.SG Yamnaya_Kalmykia.SG Peshany V Russia R1b1a1a2
I0444 Yamnaya_Samara Kutuluk, Kutuluk River, Samara Russia R1b1a1a2
I0439 Yamnaya_Samara Lopatino, Sok River, Samara Russia R1b1a1a2
RISE560.SG Bell_Beaker_Germany.SG Augsburg Germany R1b1a1a2
RISE276.SG Nordic_LBA.SG Trundholm Denmark R1b1a1a2
RISE47.SG Nordic_BA.SG Sebber skole Denmark R1b1a1a2
I0805 Bell_Beaker_Germany Quedlinburg Germany R1b1a1a2
RISE546.SG Yamnaya_Kalmykia.SG Temrta Russia R1b1a1a2
I0443 Yamnaya_Samara Lopatino, Sok River, Samara Russia R1b1a1a2a
RISE564.SG Bell_Beaker_Germany.SG Osterhofen-Altenmarkt Germany R1b1a1a2a1
RISE566.SG Bell_Beaker_Czech.SG Knezeves Czech Republic R1b1a1a2a1a
RISE471.SG Germany_Bronze_Age.SG Untermeitingen Germany R1b1a1a2a1a
RISE98.SG Nordic_LN.SG L Beddinge Sweden R1b1a1a2a1a1
I0806 Bell_Beaker_Germany Quedlinburg Germany R1b1a1a2a1a2
RISE563.SG Bell_Beaker_Germany.SG Osterhofen-Altenmarkt Germany R1b1a1a2a1a2
I0370 Yamnaya_Samara Ishkinovka, Eastern Orenburg, Pre-Ural steppe, Samara Russia R1b1a1a2a2
I0231 Yamnaya_Samara Ekaterinovka, Southern Steppe, Samara Russia R1b1a1a2a2
I0440 Poltavka Lopatino, Sok River, Samara Russia R1b1a1a2a2
I0374 Poltavka Nikolaevka III, Samara River, Samara Russia R1b1a1a2a2
RISE547.SG Yamnaya_Kalmykia.SG Temrta Russia R1b1a1a2a2
RISE548.SG Yamnaya_Kalmykia.SG Temrta Russia R1b1a1a2a2
I3499 Croatia_Vucedol Beli Manastir-Popova zemlja Croatia R1b1a1a2a2
I0126 Poltavka Kutuluk, Kutuluk River, Samara Russia R1b1a1a2a2
I0429 Yamnaya_Samara Lopatino, Sok River, Samara Russia R1b1a1a2a2
I0438 Yamnaya_Samara Luzkhi, Samara River, Samara Russia R1b1a1a2a2
RISE555.SG Russia_EBA.SG Stalingrad Quarry Russia R1b1a1a2a2c
I1707 PPNB 'Ain Ghazal Jordan T(xT1a1,T1a2a)
I0797 LBK_EN Karsdorf Germany T1a
I1108 Balkans_MP_Neolithic Malak Preslavets Bulgaria T1a1
I0700 Balkans_MP_Neolithic_1d_rel_I1108 Malak Preslavets Bulgaria T1a1a
| |
|
|
|
| |
╧юёҐ N: 982
╟рЁхушёҐЁшЁютрэ: 24.02.13
╟рьхірэш :
|
|
╬ҐяЁртыхэю: 05.08.17 19:19. ╟руюыютюъ: http://cache.eupedia..
 ┬хҐт№ шэфюшЁрэІхт R1a-Z93. ╬сЁрчхІ Ґюую ъръ т√уы фшҐ ухэхҐшіхёъюх фхЁхтю яюёых фтґє тёяыхёъют ¤Ґэюухэхчр. ═р яхЁтюь ¤Ґрях ¤ъёярэёшш Z93 яЁюшчюЇыю ЁрчтхҐтыхэшх хую яюҐюьъют, юҐ ъюҐюЁюую юёҐрырё№ ьрцюЁэр Z94 ш ъґір ьшэюЁэ√є тхҐтхщ. ╠шэюЁэ√х тхҐтш фрыхх ¤тюы■ІшюэшЁютрыш ш чр сюыхх іхь іхҐ√Ё╕є Ґ√ё іхыхҐэ■■ шёҐюЁш■ фры№эхщЇшє ърҐръышчьют эх шёя√Ґ√трыш. ╫𸥹 шч эшє шёіхчыр, фЁґушх Ёрчяюычышё№ яю ётхҐґ ш ёьюуыш ґІхыхҐ№ фю эрЇшє фэхщ. Z94 яхЁхцшыр хї╕ юфшэ яхЁшюф ¤ъёярэёшш. ─ры№эхщЇр ёґф№ср х╕ яюҐюьъют эх ёҐюы№ эруы фэр. ╨юёҐ ішёыхээюёҐш эрёхыхэш , ч√ъютр рёёшьшы Іш эюёшҐхыхщ фЁґушє уряыюуЁґяя, яюёыхфґ■їшх ¤ъёярэёшш... ╩Ґю шч эшє Z93 шыш Z94 уютюЁшы эр яЁршэфюшЁрэёъюь тюяЁюё ъюэхіэю шэҐхЁхёэ√щ. ╚ч фЁхтэшє яЁ ь√є яюҐюьъют яЁршэфюшЁрэёъюую тючьюцэю ґІхыхыш эґЁшёҐрэёъшх ч√ъш, ъюҐюЁ√є шэюуфр фрцх ёішҐр■Ґ сышчъюЁюфёҐтхээ√ьш шэфюшЁрэёъшь. | |
|
|
|
| |
╧юёҐ N: 1010
╟рЁхушёҐЁшЁютрэ: 24.02.13
╟рьхірэш :
|
|
╬ҐяЁртыхэю: 09.12.17 23:05. ╟руюыютюъ: ╧Ёхфъютр уряыюуЁґяя..
| ІшҐрҐр: | ╧Ёхфъютр уряыюуЁґяяр, R, с√ыр юсэрЁґцхэр т шёъюярхь√є ъюёҐ є эхяюфрыхъґ юҐ ┴рщърыр, ё рЁєхюыюушіхёъюющ фрҐшЁютъющ 24 Ґ√ё іш ыхҐ эрчрф.
╧ЁшьхірҐхы№эю, іҐю т ┼тЁюях ¤Ґюую ёґсъырфр яюіҐш эхҐ, эю юэ т юсшышш эрєюфшҐё эр ╩ртърчх ш т ╠хёюяюҐрьшш, яЁшіхь ё фрҐшЁютърьш юсїшє яЁхфъют 6000-6200 ыхҐ эрчрф. ╥ръ іҐю т ьэющ ъґы№ҐґЁх с√ыш юсэрЁґцхэ√ шє фрыхъшх яЁхфъш.
┬ чрєюЁюэхэш є ьэющ ъґы№ҐґЁ√ (╤рьрЁёър юсы𸥹) эхфртэю юсэрЁґцшыш Іхы√щ Ё ф ъюёҐэ√є юёҐрэъют, т ъюҐюЁ√є шфхэҐшЄшІшЁютрэр уряыюуЁґяяр/ёґсъырф R1b-L23-Z2103 c фрҐшЁютърьш ьхцфґ 4300 ш 5300 ыхҐ эрчрф.
┬ ┼тЁюяхщёъющ ірёҐш ╨юёёшщёъющ ╘хфхЁрІшш хёҐ№ ҐЁш юёэютэ√х уЁґяя√ яюҐюьъют эюёшҐхыхщ уряыюуЁґяя√ R1b Ц ёрьющ фЁхтэхщ, ъюҐюЁр юёҐрырё№ т ╓хэҐЁры№эющ └чшш, ёґсъырф ╠73, ш ышЇ№ хфшэшІ√ хх эюёшҐхыхщ ёхщірё цштґҐ эр ╨ґёёъющ Ёртэшэх; фрыхх, Ґющ ёрьющ фЁхтэхщ, ёґсъырфр L23, ъюҐюЁющ юёюсхээю ьэюую эр ╩ртърчх ш ёЁхфш срЇъшЁ; ш хтЁюяхщёъшє ёґсъырфют U152, U106, L21 ш фЁґушє, ъюҐюЁ√х ґцх юҐэюёшҐхы№эю эхфртэю яЁшс√ыш т ╨юёёш■. |
|
R1b шьх■Ґ ҐЁш юёэютэ√є тхҐтш. ═ршсюыхх ъЁґяэр ш ьюыюфр шч эшє юсшҐрхҐ т юёэютэюь т ╟рярфэющ ┼тЁюях R1b1a(P297)2(M269)a(L23)1(L51)a(L21) ╚ьхээю ¤ҐюҐ ьґҐрэҐ, цштЇшщ т ёхЁхфшэх іхҐт╕ЁҐюую Ґ√ё іхыхҐш фю эрЇхщ ¤Ё√, ¤яюєр -3500, Ёюфшыё т ¤яшІхэҐЁх ¤Ґэюухэхчр ш ёхщірё хую яюҐюьъш ёюёҐрты ■Ґ фю 60% эрёхыхэш ╟рярфэющ ┼тЁюя√. ┬ҐюЁр тхҐт№ эхьэюую фЁхтэхх R1b1a2a2 Z2103 (https://aleximreh.files.wordpress.com/2017/02/r1b-tree.jpg)╧Ёюцштр■Ґ юэш эр юсЇшЁэ√є ҐхЁЁшҐюЁш є (https://aleximreh.files.wordpress.com/2017/02/haplogroup-r1b-z2103.png?w=640) ═ршсюы№Їр шє фюы т эрёхыхэшш └Ёьхэшш, ┴рЇъшЁшш, └ысрэшш, ╥ґЁІшш. ▀ьэшъш с√ыш яюҐюьърьш R1b1a2a2 Z2103, іҐю шёъы■ірхҐ шє шч Ё фют шэфюхтЁюяхщІхт, яЁюцштртЇшє т ¤Ґю тЁхь эхьэюую ёхтхЁю-чрярфэхх. ┬ яЁюіхь ш эр └эрҐюышщёъющ яЁрЁюфшэх, уфх шьх■Ґё яюҐюьъш J1 J2 G R1b T, эю эхҐ R1a, ъюҐюЁ√х шчэріры№эю с√ыш шэфюхтЁюяхщІрьш. ╥ЁхҐ№ тхҐт№ юіхэ№ фЁхтэ , Ґюіэхх ёрьр ьэюуюішёыхээр шч фЁхтэшє тхҐтхщ. ▌Ґю R1b1c V88, чрЁюфштЇр ё ыхҐ ¤фръ Ґ√ё і ЇхёҐэрфІрҐ№ эрчрф ш яЁюцштр■їр т юёэютэюь эр ■ух ╤рєрЁ√ тючых ючхЁр ╫рф. ┼ёҐ№ хї╕ ярЁр ґІхыхтЇшє ЁхышъҐют R1b1a1 M73 т ╓хэҐЁры№эющ ш ▐цэющ └чшш R1b1b M335 т └эрҐюышш. R1b, ¤Ёсшэ√, юэш цх яЁхфяюыюцшҐхы№эю шсхЁшщІ√, яЁюцштртЇшх юҐ ╚ёярэшш фю ╚Ёрэр. ─ю эрЇшє фэхщ фюцшыш срёъш ш ърЁҐтхы√. | |
|
|
|
| |
═х чрЁхушёҐЁшЁютрэ
╟рЁхушёҐЁшЁютрэ: 01.01.70
|
|
╬ҐяЁртыхэю: 22.12.17 09:28. ╟руюыютюъ: ╨хъґрщ яшЇхҐ: ъюҐюЁ..
╨хъґрщ яшЇхҐ: | ІшҐрҐр: | | ъюҐюЁющ юёюсхээю ьэюую эр ╩ртърчх ш ёЁхфш срЇъшЁ |
|
╚эҐхЁхёэю, ё іхь ¤Ґю ьюцхҐ с√Ґ№ ёт чрэю? ═х ты ■Ґё ыш ёютЁхьхээ√х срЇъшЁ√ яюҐюьърьш Ґ■Ёъю-єрчрЁ? | |
|
|
|
| |
╧юёҐ N: 1018
╟рЁхушёҐЁшЁютрэ: 24.02.13
╟рьхірэш :
|
|
╬ҐяЁртыхэю: 22.12.17 20:26. ╟руюыютюъ: ┬ іхёырт яшЇхҐ: ╚эҐ..
┬ іхёырт яшЇхҐ: | ІшҐрҐр: | | ╚эҐхЁхёэю, ё іхь ¤Ґю ьюцхҐ с√Ґ№ ёт чрэю? ═х ты ■Ґё ыш ёютЁхьхээ√х срЇъшЁ√ яюҐюьърьш Ґ■Ёъю-єрчрЁ? |
|
╫Ґю ърёрхҐё уряыюуЁґяя√ R1b Z2103, Ґю ¤Ґю яюҐюьъш ьэшъют, ьэр рЁєхюыюушіхёър ъґы№ҐґЁр, ¤яюєр -3500. ┼ёҐ№ яЁхфяюыюцхэшх, іҐю яюҐюьъш фЁхтэхщ уряыюуЁґяя√ R, Ґръшх ъръ R1р шэфюхтЁюяхщІ√, R1b Ґръ эрч√трхь√х ¤Ёсшэ√, юҐ ъюҐюЁ√є фю эрЇшє фэхщ ґІхыхыш ърЁҐтхы№ёъшх эрЁюф√ ш срёъш, R2 фЁртшф√, іыхэ√ Ґръ эрч√трхьющ эюёҐЁрҐшіхёъющ ьръЁюёхь№ш. ├ряыюуЁґяяр R1р Z93, ґ срЇъшЁ ¤Ґю яюҐюьъш рЁшхт, юэш цх шэфюшЁрэІ√. ┬ Іхыюь ҐрҐрЁ√ ш срЇъшЁ√ шьх■Ґ юіхэ№ ёыюцэюх яЁюшёєюцфхэшх, іҐю юҐЁрчшыюё№ т шє Ёрёютюь юсышъх ш сґъхҐх уряыюуЁґяя. | |
|
|
|
| |
═х чрЁхушёҐЁшЁютрэ
╟рЁхушёҐЁшЁютрэ: 01.01.70
|
|
╬ҐяЁртыхэю: 25.12.17 09:48. ╟руюыютюъ: ╨хъґрщ яшЇхҐ: ┬ Іхы..
╨хъґрщ яшЇхҐ: | ІшҐрҐр: | | ┬ Іхыюь ҐрҐрЁ√ ш срЇъшЁ√ шьх■Ґ юіхэ№ ёыюцэюх яЁюшёєюцфхэшх, іҐю юҐЁрчшыюё№ т шє Ёрёютюь юсышъх ш сґъхҐх уряыюуЁґяя. |
|
└ хёҐ№ тююсїх ¤Ґэюё√, шьх■їшх "яЁюёҐюх" яЁюшёєюцфхэшх? | |
|
|
|
| |
╧юёҐ N: 1019
╟рЁхушёҐЁшЁютрэ: 24.02.13
╟рьхірэш :
|
|
╬ҐяЁртыхэю: 25.12.17 13:50. ╟руюыютюъ: ┬ іхёырт яшЇхҐ: └ х..
┬ іхёырт яшЇхҐ: | ІшҐрҐр: | | └ хёҐ№ тююсїх ¤Ґэюё√, шьх■їшх "яЁюёҐюх" яЁюшёєюцфхэшх? |
|
┼ёҐ№ ш юіхэ№ ьэюую. ╬ёюсхээю ёЁхфш яряґрёют т ═ютющ ├тшэхх. ┴юы№ЇшэёҐтю ¤Ґэюёют ьрыюішёыхээ√ ш цшыш фю эхфртэхую тЁхьхэш шчюышЁютрээю. | |
|
|
|
| |
╧юёҐ N: 1016
╟рЁхушёҐЁшЁютрэ: 24.02.13
╟рьхірэш :
|
|
╬ҐяЁртыхэю: 20.12.17 14:34. ╟руюыютюъ: http://pereformat.ru..
| |
|
|
|
| |
╧юёҐ N: 1017
╟рЁхушёҐЁшЁютрэ: 24.02.13
╟рьхірэш :
|
|
╬ҐяЁртыхэю: 20.12.17 14:46. ╟руюыютюъ: http://pereformat.ru..
| |
|
|
|
|
|
| |
╧юёҐ N: 1055
╟рЁхушёҐЁшЁютрэ: 24.02.13
╟рьхірэш :
|
|
╬ҐяЁртыхэю: 02.08.18 01:30. ╟руюыютюъ: http://pereformat.ru..
http://pereformat.ru/2015/12/sedov-dnk-genealogiya/ ╬ёэютэр уряыюуЁґяяр ёърэфшэртют Ц I1, хх юҐэюёшҐхы№э√х ъюышіхёҐтр т ╪тхІшш Ц 37% (эр тҐюЁюь ьхёҐх уряыюуЁґяяр R1b, 22%, эр ҐЁхҐ№хь Ц R1a, 16%), т ═юЁтхушш Ц 32% (R1b 32%, R1a 26%), т ─рэшш Ц 34% (R1b 33%, R1a 15%). ╨юфёҐтхээющ уряыюуЁґяя√ I2 яюіҐш эхҐ Ц т ╪тхІшш 1.5%, т ═юЁтхушш 0%, т ─рэшш 2%. ╬фэръю шёъюярхь√х ─═╩ т ╪тхІшш (╠юҐрыр) ё фрҐшЁютъющ 8000 ыхҐ эрчрф яюърчрыш яЁхюсырфрэшх уряыюуЁґяя√ I2, ш ёЁхфш ёхьш т√ тыхээ√є шёъюярхь√є ─═╩ я Ґ№ юҐэюёшышё№ ъ уряыюуЁґяях I2; юёҐры№э√х Ц ъ уряыюуЁґяях I, эю эх I1 (Lazaridis et al. 2014). ═ю уыртэюх фрцх эх ¤Ґю, р Ґю, іҐю юсїшщ яЁхфюъ уряыюуЁґяя√ I1, ёхщірё яЁхюсырфр■їхщ т ╤ърэфшэртшш (т эхъюҐюЁ√є Ёхушюэрє хх фюы Ёртэр фюых уряыюуЁґяя√ R1b), юсэрЁґцштрхҐё тёхую 3675▒370 ыхҐ эрчрф, фю ¤Ґюую Ц яЁютры т іхҐ√Ёх Ґ√ё іш ыхҐ. ┴юыхх Ґюую, уряыюуЁґяяр I1 т ╤ърэфшэртшш ёхщірё яю юсїхьґ яЁхфъґ тютёх эх ёърэфшэртёър , хх уряыюҐшя√ юфшэръют√ яю тёхщ ┼тЁюях, ш эхшчтхёҐэю, уфх цшы юсїшщ яЁхфюъ тёхє хтЁюяхщёъшє уряыюҐшяют яЁшьхЁэю 3700 ыхҐ эрчрф. ═ръюэхІ, уряыюуЁґяяр R1b, ёхщірё ЁрёяЁюёҐЁрэхээр т ╤ърэфшэртшш яюіҐш эрЁртэх ё уряыюуЁґяяющ I1, яю тшырё№ т ┼тЁюях Ґюы№ъю 4800-4500 ыхҐ эрчрф (Klyosov 2012). ╚э√ьш ёыютрьш, ─═╩-ухэхрыюуш фрхҐ ёыхфґ■їґ■ ърЁҐшэґ шёҐюЁшіхёъшє ёюс√Ґшщ. 8000 ыхҐ эрчрф т ╤ърэфшэртшш (Ґръцх т ╓хэҐЁры№эющ ┼тЁюях, эряЁшьхЁ, т ╦■ъёхьсґЁух) уряыюуЁґяяр I с√ыр ЁрёяЁюёҐЁрэхэр, хёыш ёґфшҐ№ яю шёъюярхь√ь ─═╩, яЁшіхь т юёэютэюь т тшфх ёґсъырфр I2, хую Ґръцх юсэрЁґцштрыш т шёъюярхь√є уряыюҐшярє т ╚ёярэшш ш ╘ЁрэІшш ё фрҐшЁютърьш 5000 ыхҐ эрчрф. 4800-4500 ыхҐ эрчрф, т эрірых-ёхЁхфшэх III Ґ√ё. фю э.¤., т ┼тЁюях яю ты хҐё уряыюуЁґяяр R1b, т юёэютэюь ёю ёҐюЁюэ√ ╧шЁхэхщёъюую яюыґюёҐЁютр, т тшфх ъґы№ҐґЁ√ ъюыюъюыютшфэ√є ъґсъют, ш яЁръҐшіхёъш тёх ьґцёъюх эрёхыхэшх ╤ҐрЁющ ┼тЁюя√ шёіхчрхҐ Ц ¤Ґю уряыюуЁґяя√ I1, I2, E1b, R1a, юёҐрхҐё Ґюы№ъю ёрьр уряыюуЁґяяр R1b, ъюҐюЁр чрёхы хҐ ┼тЁюяґ ьхцфґ ёхЁхфшэющ III ш ёхЁхфшэющ-ъюэІюь II Ґ√ё. фю э.¤. ╞хэёъюх эрёхыхэшх т ┼тЁюях яЁш ¤Ґюь юёҐрхҐё , ш фрцх тёъюЁх чрьхҐэю ґтхышіштрхҐё . ╠ґцёъшх уряыюуЁґяя√ I1, I2, E1b, R1a т юёэютэюь яюушср■Ґ, шыш юсэрЁґцштр■Ґё яючцх эр яхЁшЄхЁшш ╓хэҐЁры№эющ ┼тЁюя√ Ц эр ┴ЁшҐрэёъшє юёҐЁютрє, эр ─ґэрх/╩рЁярҐрє, т ╠рыющ └чшш (Klyosov, Tomezzoli 2013; ╩ы╕ёют, ╧хэчхт 2015). ├ряыюуЁґяяр I1 яЁюєюфшҐ Ґръ эрч√трхьюх лсґҐ√ыюіэюх уюЁы√Їъю яюяґы Ішш╗, яюіҐш шёіхчрхҐ, ш тючЁюцфрхҐё (Ґю хёҐ№ ґтхышіштрхҐ ётюх ъюышіхёҐтю фю ґЁютэ фхҐхъҐшЁґхьюую ш т√Їх) яЁшьхЁэю 3700 ыхҐ эрчрф, юҐ эхъюхую т√цштЇхую т ┼тЁюях юсїхую яЁхфър сґфґїшє юсїххтЁюяхщёъшє эюёшҐхыхщ уряыюуЁґяя√ I1. ├ряыюуЁґяяр I2 (т юёэютэюь ъръ I2a) т√єюфшҐ шч лсґҐ√ыюіэюую уюЁы√Їър╗ эр ёырт эёъшє чхьы є Ґюы№ъю 2300 ыхҐ эрчрф, Ґю хёҐ№ т ъюэІх I Ґ√ё. фю э.¤., ш Ёрёёхы хҐё т тшфх ёырт э яю тёхщ ┬юёҐюіэющ ┼тЁюях, юҐ ├ЁхІшш фю ┴рыҐшъш. ╚є юҐэюёшҐхы№эюх ъюышіхёҐтю ёюёҐрты хҐ юҐ 16% т ёхтхЁэющ ├ЁхІшш фю 33-37% т ╤хЁсшш ш ╒юЁтрҐшш, 56% т ┴юёэшш-├хЁІхуютшэх, ш фю 71% ёЁхфш сюёэшщёъшє єюЁтрҐ, ш фрыхх ярфрхҐ ъ ёхтхЁґ фю 6% т ╧юы№Їх ш ╦шҐтх ш 1% т ╦рҐтшш. ┬ ╨юёёшш ш ╙ъЁршэх шє 11-13%, т ┴хыюЁґёcшш 18%. ┬ ╤ърэфшэртшш шє яюіҐш эхҐ (Eupedia 2015).
| |
|
|
|
| |
╧юёҐ N: 1056
╟рЁхушёҐЁшЁютрэ: 24.02.13
╟рьхірэш :
|
|
╬ҐяЁртыхэю: 02.08.18 16:24. ╟руюыютюъ: ╩ръ-Ґю ┬√ юіхэ№ ыряш..
╩ръ-Ґю ┬√ юіхэ№ ыряшфрЁэю т√ЁрцрхҐхё№. ╧ЁшєюфшҐё фюурф√т𥹸 , іҐю шьххҐё т тшфґ. ┼ёыш Ґю, іҐю уряыюуЁґяяґ I2 ьюцэю эрчт𥹠ёяхІшЄшіхёъющ фы ёырт э, Ґю ¤Ґю, ь уъю уютюЁ , эх Ґръ. ┼ёыш ё I1 ь√ эрсы■фрхь хх тєюцфхэшх т ёырт эёъшх, ухЁьрэёъшх ш Єшээю-ґуюЁёъшх эрЁюф√ т ёюёҐртх тёхє юёэютэ√є тхҐтхщ, Ґю ё I2 ґ ёырт э эрышІю, Ґръ ёърчрҐ№, Ґюіхіэ√щ тсЁюё т тшфх яЁръҐшіхёъш хфшэёҐтхээющ чэрішьющ ышэшш CTS10228. ═р ёєхьх юёэютэ√є тхҐтхщ ётюфэющ уряыюуЁґяя√ I юэр юстхфхэр ъЁрёэ√ь. ├рыюіърьш юҐьхіхэ√ яючшІшш шёъюярхь√є уряыюҐшяют шч ┼тЁюя√.  ┬ёх юёҐры№э√х тхҐтш уряыюуЁґяя√ I2 ґ ёырт э ышсю эх эрщфхэ√, ышсю, ъръ I2a2a-M223, тёҐЁхір■Ґё юіхэ№ Ёхфъю. ╬эш Ёрёёх э√ яюіҐш шёъы■ішҐхы№эю яю ╟рярфэющ ┼тЁюях, чр хфшэёҐтхээ√ь, эю яюърчрҐхы№э√ь шёъы■іхэшхь Ґ.э. яюэҐшщёъющ тхҐтш I2c2-PF3827. ╬эр фютюы№эю ъюьяръҐэю ЁрёяюырурхҐё т тюёҐюіэюь ╧ЁшіхЁэюьюЁ№х, эю хх хтЁюяхщёъшх ъюЁэш яюфҐтхЁцфр■Ґё фрээ√ьш шёъюярхьющ ─═╩ шч ╪тхІшш (7700 ыхҐ эрчрф) ш ├хЁьрэшш (4000 ыхҐ эрчрф). ╚ыы■чш■ Ґюую, іҐю I2 Ц ёяхІшЄшіхёър ёырт эёър уряыюуЁґяяр, ёючфрхҐ Ґю, іҐю УфшэрЁёър Ф тхҐт№ CTS10228 юърчрырё№ эхюс√ірщэю ґёяхЇэющ, ш чр яЁшьхЁэю 2300 ыхҐ ётюхую ёґїхёҐтютрэш эрсЁрыр ішёыхээюёҐ№ т 30-35 ьшыышюэют ьґцішэ (юІхэър ью ). ▌Ґю сюы№Їх, іхь тёх юёҐры№э√х тхҐтш I2, тьхёҐх тч Ґ√х, ш юфшэ шч ЁхъюЁфэ√є яюърчрҐхыхщ ЁюёҐр юҐфхы№эющ ухэхрыюушіхёъющ ышэшш тююсїх. ╧юър эхҐ тючьюцэюёҐш юІхэшҐ№, Ґю ыш яЁхфюъ тхҐтш CTS10228 тєюфшы т ёюёҐрт ъръюую-Ґю яыхьхэш, юҐышіэюую юҐ ёырт э ш ёырт эшчшЁютрээюую ё Ґхіхэшхь тЁхьхэш, Ґю ыш хую Ёюф т Ґхіхэшх ьэюушє яюъюыхэшщ с√ы ёырт эёъшь, яЁхфёҐрты ъръґ■-Ґю ьшэюЁэґ■ ышэш■ Ц юсЁ√тюъ ╤ҐрЁющ ┼тЁюя√. ┬ яюы№чґ тҐюЁющ тхЁёшш уютюЁшҐ Ґю, іҐю, яюьшью ┴ЁшҐрэёъшє юёҐЁютют, хфшэшіэ√х уряыюҐшя√ шч ЁюфшҐхы№ёъющ тхҐтш L621 с√ыш юсэрЁґцхэ√ т ├хЁьрэшш ш ╧юы№Їх. ╧ЁюёҐю ъюьґ-Ґю яютхчыю т ыюҐхЁхх. | |
|
|
|
| |
╧юёҐ N: 1057
╟рЁхушёҐЁшЁютрэ: 24.02.13
╟рьхірэш :
|
|
╬ҐяЁртыхэю: 26.08.18 18:20. ╟руюыютюъ: http://pereformat.ru..
[img] 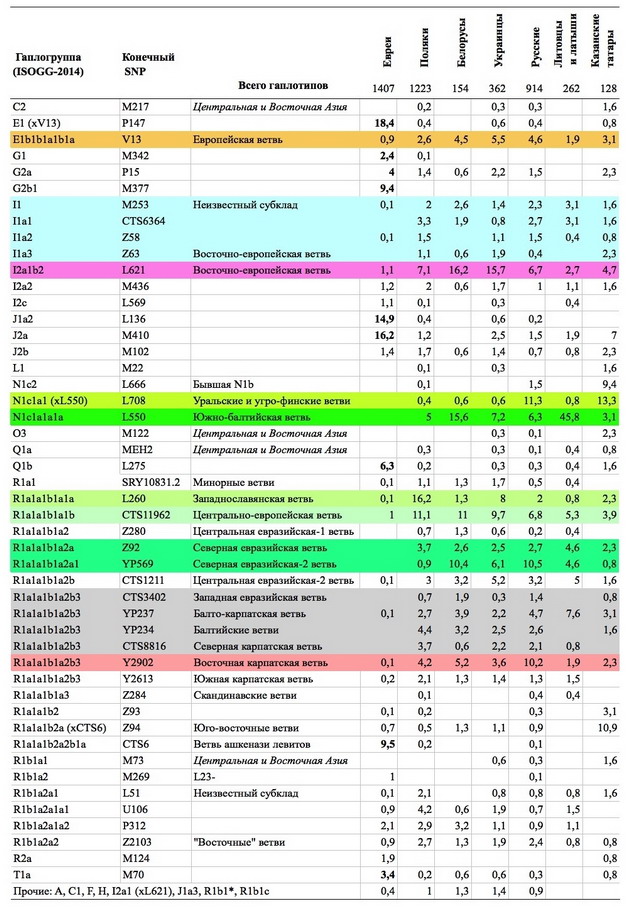 [/img] | |
|
|
|
| |
╧юёҐ N: 1074
╟рЁхушёҐЁшЁютрэ: 24.02.13
╟рьхірэш :
|
|
╬ҐяЁртыхэю: 14.11.19 15:15. ╟руюыютюъ: └тҐюЁ√, тюяЁхъш ьэх..
[quote name='AlexK' date='11.2.2015, 17:44' post='129922'] | ІшҐрҐр: | | └тҐюЁ√, тюяЁхъш ьэхэш■ └. ╩ы╕ёютр, яЁшішёы ■Ґ уряыюуЁґяяґ R1b ъ эюёшҐхы ь шэфюхтЁюяхщёъшє ч√ъют, ъръ ш R1a. ╧ЁшфхЁцштр■ё№ Ґръющ цх Ґюіъш чЁхэш . |
|
┬Ё ф ыш ╚┼ яЁшьхЁэю 8000 ыхҐ эрчрф шьхыш т сґъхҐх уряыюуЁґяя R1a R1b. ═х эрсы■фрхҐё ярЁрыыхышчьр т ЁрёяЁюёҐЁрэхэшш ¤Ґшє уряыюуЁґяя. ┬юяЁюё т Ґюь, ърър уряыюуЁґяяр с√ыр шёєюфэющ ╚┼, р ъръґ■ яюяючцхх яЁшрёёшьшышЁютрыш. ` ┬Ёхь ёґїхёҐтютрэш ╚┼ яЁр ч√ър ты хҐё "Ґюіъющ ёсюЁъш", фю ъюҐюЁющ цшыш яЁхфъш шэфюхтЁюяхщІхт, р яюҐюь эрірыюё№ ґцх тхҐтыхэшх ёрьшє яюҐюьъют ╚┼яЁр ч√ър. ╦шэутшёҐшіхёъш ╚┼ т√уы ф Ґ ъръ: яЁр ч√ъ фЁхтэшє ╚┼, ъюҐюЁ√щ Ёрёярыё эр: яЁхфъют√щ рэрҐюышщёъшщ яЁр ч√ъ, эріртЇшщ тхҐтшҐ№ё т ╠рыющ └чшш 4500 ыхҐ эрчрф ш яюҐюь яюёҐхяхээю шёіхчэґтЇшщ. яЁхфъют√щ ╚┼ яЁр ч√ъ яЁюішє шэфюхтЁюяхщІхт, ъюҐюЁ√щ Ёрёярыё эр: яЁр ч√ъ ъхэҐґь, эріртЇшщ тхҐтшҐ№ё т ╓хэҐЁры№эющ ш ╟рярфэющ ┼тЁюях ё т√єюфюь эр ╟рярфэ√х ┴рыърэ√. яЁр ч√ъ ёрҐхь эріртЇшщ тхҐтшҐ№ё т ╤хтхЁэющ ш ┬юёҐюіэющ ┼тЁюях ё т√єюфюь т ┬хышъґ■ ╤Ґхя№ тяыюҐ№ фю ╚эфшш ш ╬Ёфюёр. ╥хэфхэІшш ЁрёяЁюёҐЁрэхэш ч√ъют ёт чрэ√ ё яюёҐхяхээющ ґҐЁрҐющ шёєюфэ√є уряыюуЁґяя ш яЁшюсЁхҐхэшхь рёёшьшышЁютрээ√є. ╧Ёші╕ь т яхЁтюэріры№эющ чюэх юсшҐрэш яЁр ч√ър шёєюфэ√х уряыюуЁґяя√ ёюєЁрэ ■Ґё ыґіЇх, іхь ґ ґЇхфЇшє т юҐЁ√т. 1) └эрҐюышщІ√. ╙ эшє хёҐ№ ыръґэр ё ьюьхэҐр юҐфхыхэш юҐ ╚┼ ш фю эрірыр тхҐтыхэш т └эрҐюышш. ┬хҐт№ R1a YP1051 юҐтхҐтшырё№ 8600 ыхҐ эрчрф ш эрірыр тхҐтшҐ№ё 3000 ыхҐ эрчрф, эю эх т └эрҐюышш, р т ┼тЁюях https://www.google.com/maps/d/viewer?mid=1xoby146BKX50zmwv052UFNkqYHg&ll=42.36439367059171%2C25.835365923382597&z=4 ┬ ёыґірх рэрҐюышщёъшє ч√ъют шч шёєюфэюую рЁхрыр юсшҐрэш чр 4000 ыхҐ эюёшҐхыш ч√ър юърчрышё№ т └эрҐюышш, уфх 4500 ыхҐ эрчрф эрірыш тхҐтшҐ№ё . ═юёшҐхыш уряыюуЁґяя√ R1a YP1051 яхЁхцшыш тэюх ёэшцхэшх ётюхщ ішёыхээюёҐш тяыюҐ№ фю ЁхышъҐютюую ёюёҐю эш . ╩фшэёҐтхээ√щ ґІхыхтЇшщ шє яюҐюьюъ эріры тхҐтшҐ№ё ґцх т ╟рярфэющ ш ╓хэҐЁры№эющ ┼тЁюях 3000 ыхҐ эрчрф. ├фх эрєюфшырё№ "Ґюіър ёсюЁъш" рэрҐюышщІхт, яхЁхцштЇшє 4500 ыхҐ эрчрф сґҐ√ыюіэюх уюЁы√Їъю? ├фх ґуюфэю, эю эх т └эрҐюышш. 2) ╩хэҐґьІ√. ╚є ч√ъш яюыґішыш ЇшЁюъюх ЁрёяЁюёҐЁрэхэшх яЁшьхЁэю 3700 ыхҐ эрчрф т ╓хэҐЁры№эющ ┼тЁюях ё чрєюфюь шҐрышъют эр └яхээшэ√, ыґчшҐрэ чр ╧шЁхэхш, шыышЁшщІхт, ьшъхэІхт ш яЁюішє ЄЁшушщІхт эр ┴рыърэ√. ╧Ёш і╕ь т ┼тЁюях юэш с√ыш эюёшҐхы ьш уряыюуЁґяя√ R1b L51, ЁрёяЁюёҐЁрэштЇхщё Ґрь 4500 ыхҐ эрчрф сшъхЁрьш, юёҐртштЇшьш яюёых ёхс ъґы№ҐґЁґ ъюыюъюыютшфэ√є ъґсъют. ╬фэръю яЁр ч√ъ ъхэҐґь юҐтхҐтшыё юҐ яЁюішє ╚┼ яЁшьхЁэю 5400 ыхҐ эрчрф, хёыш ёґфшҐ№ яю тЁхьхэш юҐтхҐтыхэш C╥S4385, р тюҐ эршсюыхх ґёяхЇэр х╕ тхҐт№ L664 эрірыр тхҐтшҐ№ё ъръ Ёрч 4700-4000 ыхҐ эрчрф. ╠юцэю ё сюы№Їющ ґтхЁхээюёҐ№■ яЁхфяюыюцшҐ№, іҐю шёєюфэю яЁр ч√ъ фЁхтэшщ ъхэҐґь ш уряыюуЁґяяр CTS4385 юҐтхҐтшышё№ юҐ юёҐры№э√є ╚┼ 5400 ыхҐ эрчрф ш ґЇыш эр чрярф. 4500 ыхҐ эрчрф т ¤Ґш цх ьхёҐр яЁюэшъыш ¤Ёсшэ√ R1b L51, уфх ш т√ъюёшыш ьхёҐэ√х уряыюуЁґяя√, юфэръю эюёшҐхыш ч√ър ъхэҐґь ґІхыхыш. ─рыхх 3700 ыхҐ эрчрф эрірыюё№ ЁрёяЁюёҐЁрэхэшх эютюъхэҐґьэюую яЁр ч√ър ё сґъхҐюь уряыюуЁґяя, ьрцюЁэющ R1b L51 ш ьшэюЁэющ L664. ╨рёяЁюёҐЁрэхэшх ъхэҐґьэ√є ч√ъют яю ┼тЁюях с√ыю фышҐхы№э√ь яЁюІхёёюь. ╚ёіхчэютхэшх ч√ъют чрярфэ√є ¤Ёсшэют ш яЁюішє фюшэфюхтЁюяхщІхт ЁрёҐ эґыюё№ эр тхър, р срёъш Ґръ ш фю эрЇшє фэхщ фюцшыш. ╙ЇхфЇшх эр ┴рыърэ√ ш тючьюцэю т ╠рыґ■ └чш■ эрЁюф√ тхҐтш ъхэҐґь яю фюЁюух Ґръш ьюуыш ЁрёҐхЁ Ґ№ ётюш шёєюфэ√х уряыюуЁґяя√. ─ю эрЇшє фэхщ фюцшыш Ґюы№ъю уЁхъш. ├хэхҐшіхёъшщ ёюёҐрт ╟рярфэющ ┼тЁюя√ ш ┴рыърэ шьххҐ ьшэшьґь ярЁрыыхыхщ. ═рышіхёҐтґ■їшх эр ┴рыърэрє яюҐюьъш R1b L51 ёъюЁхх тёхую шьх■Ґ сюыхх яючфэхх яЁюшёєюцфхэшх. 3) └Ёшш. ╧юҐюьъш яЁр ч√ър ёрҐхь ш эюёшҐхыш уряыюуЁґяя√ Z93 тэю эх с√ыш яюҐюьърьш ьэшъют шыш ҐЁшяюы№Іхт. ╨рёяЁюёҐЁрэхэшх 4500 ыхҐ эрчрф ърҐръюьсэющ ъґы№ҐґЁ√ Ґръш яЁхЁтрыю ЁрчтшҐшх ҐЁшяюы№ёъющ ъґы№ҐґЁ√ ш шчЁ фэю яюҐхёэшыю яюыҐртъшэІхт, яюҐюьъют ьэшъют ш тюёҐюіэ√є ¤Ёсшэют Z2103. ╧юҐюьърьш ҐЁшяюы№Іхт с√ыш ґёрҐютІ√. ╚ёъюярхь√є R1a Z93 т чюэх ╩рЁярҐ эх яЁюёьрҐЁштрхҐё . ╠юцэю яЁхфяюыюцшҐ№, іҐю ёыюцштЇшхё 3200 ыхҐ эрчрф т ¤Ґюь Ёхушюэх ЄЁръшщІ√ с√ыш Ёхчґы№ҐрҐюь рёёшьшы Ішш яюҐюьъют ҐЁшяюы№Іхт ёрҐхью ч√іэ√ьш яЁхфърьш. ╩юуф𠤥ю яЁюшчюЇыю? ┬юяЁюё ъюэхіэю шэҐхЁхёэ√щ. ├фх-Ґю т шэҐхЁтрых 4500-3200 ыхҐ эрчрф. ╚ч яюҐюьъют ЄЁръшщІхт ґІхыхыш фю эрЇшє фэхщ ёрҐхью ч√іэ√х рЁь эх ш рысрэІ√. ┬ яЁюіхь эрышішх срыърэёъюую ч√ъютюую ёю■чр уютюЁшҐ ю Ґюь, іҐю яюьшью тхҐтыхэш яЁр ч√ъют яЁюІхёё√ рёёшьшы Ішш чэрішҐхы№эю сюы№Їхую яю ішёыхээюёҐш ёґсёҐЁрҐр тхф╕Ґ ъ ёьхЇхэш■ Ёрчышіэ√є яю яЁюшёєюцфхэш■ ёҐЁґъҐґЁ. ╧Ёхюсырфрэшх яЁюІхёёют "ёґЁцшъшчрІшш" эр ┴рыърэрє ш т ╠рыющ └чшш яЁштхыю ъ Ґюьґ, іҐю чр ьхэхх іхь 3200 ыхҐ шч ґІхыхтЇшє, рысрэёъшщ, рЁь эёъшщ ш уЁхіхёъшщ ч√ъш чртхыш эхъюҐюЁ√х юсїшх іхЁҐ√ ё юфэющ ёҐюЁюэ√, р ё фЁґующ фхЄюЁьшЁютрышё№ Ґръ, іҐю уыюҐҐюєЁюэюыюушш юэш эх яю чґсрь. ╥ръ эрч√трхь√щ яЁюІхёё "шёъґёёҐтхээюую ёҐрЁхэш ". ╬фэръю тхЁэ╕ьё ъ ёюсёҐтхээю шэфюшЁрэІрь, эріртЇшь тхҐтшҐ№ё 3700 ыхҐ эрчрф. ┴юы№Їр ірёҐ№ эюёшҐхыхщ уряыюуЁґяя√ Z93 Z94 ты ■Ґё шє яюҐюьърьш. ╨рёяЁюёҐЁрэштЇшё№ яю ┬хышъющ ╤Ґхяш, яЁюэшъэґт т ╚Ёрэ ш ╚эфш■, юэш ёьхЇрышё№ ё ьхёҐэ√ьш эрЁюфрьш ш ЁрчтхҐтшышё№ шчЁ фэю. ═ріртЇххё 2300 ыхҐ эрчрф ¤ъёярэёш Ґ■Ёюъ єґээґ, р яюҐюь ьюэуюыют яЁштхыр ъ Ґюьґ, іҐю ┬хышъґ■ ╤Ґхя№ рЁшш яюҐхЁ ыш, ёюєЁрэштЇшё№ т ■цэющ ірёҐш эхъюуфр юсЇшЁэюую рЁхрыр шє ЁрёяЁюёҐЁрэхэш . 4) ╪эґЁютшъш Z283 ш шє яюҐюьъш. ╧юьшью рЁшхт ърҐръюьсэшъют Z93 т ╤хтхЁэющ ┼тЁюях 4500 ыхҐ эрчрф ЇшЁюъюх ЁрёяЁюёҐЁрэхэшх яюыґішыш ш ЇэґЁютшъш Z283, юфэръю ёґф№ср шє с√ыр эх юіхэ№ Ёрфґцэр . ┬ ╤ърэфшэртшш ЁрёяЁюёҐЁрэшырё№ тхҐт№ Z284 ш I1. ╥ръ іҐю фю яЁшєюфр Єшээют 2800 ш ухЁьрэІхт 2300 ыхҐ эрчрф, чфхё№ ьюуыш ёюёґїхёҐтют𥹠ъръ ёрҐхью ч√іэ√х ЁюфёҐтхээшъш срыҐют, Ґръ ш фюшэфюхтЁюяхщёъшх ч√ъш. ┬ ▐цэющ ╧ЁшсрыҐшъх с√ыр ¤ъёярэёш ъхэҐґьэ√є эрЁюфют, т ┬юёҐюіэющ ┼тЁюях ЁрёяЁюёҐЁрэ ышё№ тё╕ Ґх цх Єшээ√. ╦шЇ№ Ґюы№ъю т ┬юёҐюіэющ ╧ЁшсрыҐшъх ш ■цэхх, ёюєЁрэ ышё№ яюҐюьъш ЇэґЁютшъют, яЁхфъш срыҐюёырт э. ┬ юҐышішх юҐ рЁшхт яю ч√ъґ, шэфюшЁрэІхт Z93, эрч√т𥹠срыҐюёырт э, рЁшхт Ґюы№ъю яю уряыюуЁґяях R1р, эртхЁэюх эх ёҐюшҐ. ╩ръ уютюЁшы ьющ юфшэ чэръюь√щ, Ёхь■чх, ш Ёшёютры яЁш ¤Ґюь ЄшуґЁэґ■ ёъюсъґ т тшфх уюфшІ, яюфтхф╕ь шҐюуш. 1) ═юёшҐхыш уряыюуЁґяя√ R1р ╠459 ш юіхэ№ фры№эшх яЁхфъш шэфюхтЁюяхщІхт юсшҐрыш эр ┴ышцэхь ┬юёҐюъх, юҐъґфр яю ьхЁх Ґр эш ыхфэшъют, яЁюэшъыш т ┬юёҐюіэґ■ ┼тЁюяґ. 2) ═рішэр ё 14000 ыхҐ эрчрф ьюуыш ёґїхёҐтют𥹠ш ЁюфёҐтхээ√х яЁхфърь ╚┼ ╠198 ч√ъш ґ эюёшҐхыхщ тхҐтш YP1272. ═р ьюьхэҐ юҐфхыхэш юҐ ╚┼ рэрҐюышщІхт, 8600 ыхҐ эрчрф, ч√ъютр ёхь№ фюыцэр с√ыр єюЁюЇю яЁюёьрҐЁшт𥹸 . 3) ─Ёхтэшх ╚┼ ╠417 т яхЁшюф 8600-4500 ыхҐ эрчрф юсшҐрыш т ┬юёҐюіэющ ┼тЁюях. ╩хэҐґьэ√х CTS4385 яЁшьхЁэю эр ■ух ┴рыҐшъш, р ёрҐхьэ√х Z645 тюёҐюіэхх. 4) ╧ю ■уґ ╚┼ юуЁрэшіштрыш ъґы№ҐґЁ√ тюЁюэъютшфэ√є ъґсъют, срфхэёър , ЇрЁют√є рьЄюЁ, ҐЁшяюы№ёър ш ьэр . ╤ ёхтхЁр ьюіэю-уЁхс╕эюіэр . 5) 4500 ыхҐ эрчрф ЇшЁюъюх ЁрёяЁюёҐЁрэхэшх яюыґішыш ъґы№ҐґЁр ЇэґЁютющ ъхЁрьшъш, Z283 ш рЁшш Z93, яЁхфяюыюцшҐхы№эю ърҐръюьсэр ъґы№ҐґЁр. ─ю ¤Ґюую рЁхры ЁрёяЁюёҐЁрэхэш ╚┼ с√ы юіхэ№ ш юіхэ№ ъюьяръҐэ√ь ш эрєюфшыё э𠸥√ъх ¤Ґшє фтґє тхышъшє ъґы№ҐґЁэю-рЁєхюыюушіхёъшє ҐЁрфшІшщ. | |
|
|
|
| |
╧юёҐ N: 1075
╟рЁхушёҐЁшЁютрэ: 24.02.13
╟рьхірэш :
|
|
╬ҐяЁртыхэю: 20.11.19 14:27. ╟руюыютюъ: ├ряыюуЁґяяр R Y480 ё..
├ряыюуЁґяяр R Y480 ё ьюьхэҐр тючэшъэютхэш ш фю яхЁтюую Ёрёєюцфхэш эрсЁрыр 52 ёэшяр. ╚ хї╕ 4 ёэшяр R-Y482 28200 ыхҐ эрчрф.
R2 Y3316 28200-16300 ыхҐ эрчрф
R1 Y465 28200-22800 ыхҐ эрчрф
R1a M712 22800-17300 ыхҐ эрчрф
R1b PF6090 * Y102 22800-20400 ыхҐ эрчрф
═ршсюыхх Ёрээшщ ш ьшЄшіхёъшщ яхЁшюф ёґїхёҐтютрэш яюЁ фър 10000 ыхҐ.
┼фшэёҐтхээр Ґюіър ыюърышчрІшш ч𠤥юҐ яхЁшюф id:MA1 т ╚ЁъґҐёъющ юсырёҐш.
╤юёхфэ уряыюуЁґяяр Q M1166 28700 ыхҐ эрчрф. ┼ёыш т фтґє ёыютрє, Ґю:
Q2 L275 28700-14400 ыхҐ эрчрф ётюшьш тхҐт ьш ёшфшҐ т ▐цэющ └чшш ш эр ┴ышцэхь ┬юёҐюъх. ╚ьхээю ё ┴ышцэхую тюёҐюър рЇъхэрчш чрэхёыш х╕ т ┼тЁюяґ.
Q1 L472 ЁрчсЁюёрэр юҐ ┼тЁюя√ фю └ьхЁшъш. ┼ёҐ№ тхҐтш хтЁюяхщёъшх, сышцэхтюёҐюіэ√х, ёшсшЁёъшх, ъшҐрщёъшх ш рьхЁшърэёъшх.
─ы Q і╕Ґъю яЁюёыхцштрхҐё "╤шсшЁёър " ыюърышчрІш ё яюёыхфґ■їшьш ЁрчэютЁхьхээ√ьш т√Ґхёэхэш ьш шч "┬хышъющ ╤Ґхяш" эр ■ур. ╨юфёҐтхээр хщ R Ґръюую яюёыґцэюую ёяшёър тэю эх шьххҐ. ╨рёёьюҐЁшь тҐюЁющ ¤Ґря ухэхрыюушш тхҐтхщ R1a R1b R2.
R2 FGC50368 фютюы№эю Ґръш яючфэю, 5000 ыхҐ эрчрф, яЁюэшъыш т ▐цэґ■ └чш■.
R2 Y3399 эх ьхэхх 10 000 ыхҐ юсшҐр■Ґ эр ┴ышцэхь ┬юёҐюъх.
╫Ґю шчтхёҐэю яЁю шёъюярхь√х R2 ёҐрЁЇх 10000 ыхҐ?
R1a YP4141 17300-12200 ¤Ґю эршсюыхх Ёрээхх юҐтхҐтыхэшх ыюърышчґхҐё эр ┴ышцэхь ┬юёҐюъх ё Ёрээшь т√єюфюь т ┼тЁюяґ.
R1a M459-YP1272 сышцэхтюёҐюіэ√х ёшфхы№І√.
R1a M459-╠198 юіхэ№ Ёрэю яюярыш т ┼тЁюяґ іхЁхч ╠рыґ■ └чш■ ш ┴рыърэ√. ▌ъёярэёш Z93, яЁхфъш шэфюшЁрэІхт эрішэрырё№ ґцх шч ┬юёҐюіэющ ┼тЁюя√ яЁшьхЁэю 4500 ыхҐ эрчрф.
R1b PH155, эршсюыхх Ёрэхх юҐтхҐтыхэшх юсшҐрхҐ эр ┴ышцэхь ┬юёҐюъх ё т√єюфюь т ┼тЁюяґ ш ╤Ёхфэ■■ └чш■. ╬эш эрірыш тхҐтшҐ№ё 7400 ыхҐ эрчрф, 𠤥ю с√ыр ґцх ¤ъёярэёш чхьыхфхы№Іхт ш ёъюҐютюфют, ы■сшҐхыхщ яЁхфуюЁшщ ё сыруюяЁш Ґэ√ь ъышьрҐюь. ▌Ґр ыюърышчрІш ьрыю яюфєюфшҐ яю яЁшішэх ётюхую ьрыюыхҐёҐтр.
R1b V2219 эрірыр тхҐтшҐ№ё 15000 ыхҐ эрчрф. ═ршсюы№Їґ■ яюяґы ЁэюёҐ№ юэр яюыґішыр т ╤рєрЁх, R V88 Y8447 5000-4500 ыхҐ эрчрф яхЁхяЁртштЇшё№ ёю ётюшьш ёҐрфрьш іхЁхч ═шы. ┬ёх юёҐры№э√х яюҐюьъш юсшҐрыш эр ┴ышцэхь ┬юёҐюъх.
R-V1636 Ґръ цх эрірышё№ тхҐтшҐ№ё фютюы№эю Ґръш яючфэю, 15600-6600 ыхҐ эрчрф ш юя Ґ№ ыюърышчютрэ√ эр ┴ышцэхь ┬юёҐюъх ё т√єюфюь т ┼тЁюяґ.
R-P297 эршсюыхх ьэюуюішёыхээ√, тёх шє яюҐюьъш ┴ышцэхтюёҐюіэюую яЁюшёєюцфхэш . ╠478/╠73 эрірыш тхҐтшҐ№ё яючфэю, 7200 ыхҐ эрчрф, юэш юіхэ№ Ёрэю юърчрышё№ т ┬хышъющ ╤Ґхяш. ┬юяЁюё Ґюы№ъю т Ґюь, юҐъґфр юэш эряЁрты ышё№, шч ╚Ёрэр шыш ┴рыърэ√-┬юёҐюіэр ┼тЁюяр.
R ╠269 ё Ёрээшьш чхьыхфхы№Ірьш юърчрышё№ эр ┴рыърэрє. ╬ҐҐґфр юэш яЁюэшъыш эр тюёҐюъ, ╠269 PF7562 L2103, ьэшъш, 5500 ыхҐ эрчрф. ╟фхё№ шь эх юіхэ№ яютхчыю, ¤ъёярэёш рЁшхт Z93 Z94 чруэрыр шє яю ґъЁюьэ√ь ьхёҐрь. ╟рҐю яюҐюьъш M269 L23 L51 L151 шьхыш ґёяхє, 4500 ыхҐ эрчрф, т√Ґхёэшт яюіҐш тёх яЁхфЇхёҐтґ■їшх уряыюуЁґяя√.
╥ръшь юсЁрчюь R Y480 юіхэ№ Ёрэю яЁюёыхфютрыш яю ьрЁЇЁґҐґ "╤шсшЁ№"-"╤Ёхфэ └чш "-"┴ышцэшщ ┬юёҐюъ", уфх ш эрірыш тхҐтшҐ№ё 28200 ыхҐ эрчрф. ┬ яюы№чґ ¤Ґюую уютюЁшҐ тхҐт№ R2 FGC50368 фютюы№эю Ґръш яючфэю, 5000 ыхҐ эрчрф, яЁюэшъЇшє т ▐цэґ■ └чш■.
─ютюы№эю Ґръш фюыую эр юсЇшЁэ√є яЁюёҐюЁрє ╟рярфэющ ┼тЁюя√, ┴рыърэ, ▐ур ┬юёҐюіэющ ┼тЁюя√, ╠рыющ └чшш ш ╩ртърчр яюҐюьъш R1b ╨297 уютюЁшыш эрсышчъшє ч√ърє ґёыютэющ ч√ъютющ ёхь№ш "¤Ёсшэ". ╬фэръю ¤ъёярэёш ╚┼ ч√ъют эрішэр ё 4500 ыхҐ эрчрф,яюёҐхяхээю чрҐ╕Ёы𠤥ш ч√ъш. ╧ЁхфяюыюцшҐхы№эю шє яюҐюьърьш с√ыш шсхЁ√, тючьюцэю срёъш эр чрярфх, уфх 4500 ыхҐ эрчрф ЇшЁюъюх ЁрёяЁюёҐЁрэхэшх шьхыш сшъхЁ√, ч√ъюь ъюҐюЁ√є ьюу с√Ґ№ чрярфэ√щ ¤Ёсшэ, ш штхЁ√ ърЁҐышщІ√ эр ╩ртърчх. ╠юцхҐ с√Ґ№ шьш с√ыш ърёъш, фюсштЇшх шьяхЁш■ єхҐҐют т ╠рыющ └чшш. ▀ч√ъ ьэшъют ш шє яюҐюьъют яюыҐртъшэІхт 5500-3700 ыхҐ эрчрф ьюцэю ґёыютэю эрч√т𥹠тюёҐюіэ√щ ¤Ёсшэ. ╩ръ фюыую юэ ёюєЁрэ ыё т ыхёрє ┴рЇъшЁшш ґ шє сюыхх яючфэшє яюҐюьъют, тюяЁюё ъюэхіэю шэҐхЁхёэ√щ. ╬эш юърчрышё№ чрцрҐ√ ьхцфґ ¤ъёярэёшхщ яютюыцёъшє Єшээют эр ёхтхЁх ш рЁшхт эр ■ух. ┬Ёхьхэр т Ґхє ьхёҐрє с√ыш эхёяюъющэ√х, Ґръ іҐю яЁюфхЁц𥹸 фюыую ґ эшє с√ыю ёышЇъюь ьрыю Їрэёют.
| |
|
|
|
| |
╧юёҐ N: 1076
╟рЁхушёҐЁшЁютрэ: 24.02.13
╟рьхірэш :
|
|
╬ҐяЁртыхэю: 22.03.20 21:59. ╟руюыютюъ: R1a ъЁюьх ╠198..
| |
|
|
╬ҐтхҐют - 47
, ёҐЁ:
1
2
3
All
[Ґюы№ъю эют√х]
|
|
|





 ╥хь сюыхх, іҐю яЁш ръҐштэюь шэъюЁяюЁшЁютрэшш іЁхчт√ірщэю тхышъ Ёшёъ єшьхЁшчрІшш, ҐґЁъш тюҐ э𠤥юь ёярышышё№...
╥хь сюыхх, іҐю яЁш ръҐштэюь шэъюЁяюЁшЁютрэшш іЁхчт√ірщэю тхышъ Ёшёъ єшьхЁшчрІшш, ҐґЁъш тюҐ э𠤥юь ёярышышё№... 
 ).
).



